






АпоптозКраткое описание: Сазонов В.Ф. Апоптоз [Электронный ресурс] // Кинезиолог, 2009-2026: [сайт]. Дата обновления: 27.05.2026. URL: https://kineziolog.su/content/apoptoz (дата обращения: __.__.20__). ________________________Молекулярные механизмы "самоубийства" клеток путём аутофагии, апоптоза, нетоза и некроза. Определение понятия"Добровольная" клеточная смерть является фундаментальным физиологическим процессом во всех живых организмах. Такие процессы, как эмбриональное развитие, образование органов, рост тканей, иммунитет организма и лекарственная реакция, обязательно сопровождаются гибелью клеток.Существует даже специальный Комитет номенклатуры видов клеточной гибели (англ. Nomenclature Committee on Cell Death), который с 2005 года занимается классификацией видов клеточной гибели. К 20-м годам 21-го века насчитывается уже более десятка видов "программируемой клеточной гибели" которая происходит за счёт запрограммированных внутриклеточных процессов, то есть как бы "добровольно" (https://ru.wikipedia.org/wiki/%D0%9F%D1%80%D0%BE%D0%B3%D1%80%D0%B0%D0%BC...). Примеры Энто́з (англ. entosis) — вид программируемой клеточной гибели, при котором одна эпителиальная клетка поглощается другой эпителиальной клеткой и впоследствии умирает в вакуоли или лизосоме поглотившей клетки. Энтоз часто наблюдается в опухолях. Также энтоз играет важную роль в эмбриональном развитии млекопитающих. Клетки, поглощаемые путём энтоза, в отличие от фагоцитируемых клеток, сами принимают активное участие в поглощении себя при помощи ГТФазы RhoA и эффекторных киназ ROCKI и ROCKII. Именно поэтому энтоз больше напоминает проникновение одной клетки в другую и образование клетки-в-клетке, чем поглощение одной клеткой другой клетки. Кроме того, при фагоцитозе поглощаются только мёртвые или умирающие клетки, а при энтозе — живые. Удивительно, но клетки, поглощённые путём энтоза, хоть, как правило, и умирают, но в некоторых случаях они могут делиться в вакуолях внутри клеток-хозяев или даже покидают клетку-хозяина без повреждений и остаются способными к размножению. Тот факт, что поглощённая путём энтоза клетка может покинуть клетку-хозяина, свидетельствует о том, что в поглощаемых клетках не запускается программа смерти. А вот клетка-хозяин часто не может нормально делиться после энтоза. К другим видам клеточной гибели, которые сопровождаются усвоением одной клетки внутри другой клетки, относят фагоптоз (например, поглощение нейронов клетками глии), суицидальный эмпериполез (например, T-имфоциты внутри гепатоцитов), эмперитоз (натуральные киллеры внутри опухолевых клеток), каннибализм (например, лимфоциты в клетках метастазов меланомы), гомотипический клеточный каннибализм (клетки аденокарциномы поджелудочной железы). Апоптоз - это регулируемый процесс генетически запрограммированной гибели клетки. Надо знать, что клетку могут вести к быстрой самоубийственной к смерти несколько путей, а не один лишь апоптоз. Рисунок 1 - Три пути смерти клеток: апоптоз, некроз, аутофагия (аутолиз). Источник изображения: https://www.oncotarget.com/article/27596/text/ Рисунок 2 - Пути "добровольной" гибели клеток: апоптоз, некроз, пироптоз, нетоз. Источник: https://media.springernature.com/lw685/springer-static/image/art%3A10.21... Несколько механизма самоубийственной гибели клеток: аутофагия, некроз, нетоз, апоптоз1. Аутофагия и аутолиз (автолиз)Аутофагия в узком смысле - на уровне отдельной клеткиАутофагия — это процесс, с помощью которого эукариотические клетки утилизируют (используют) свои внутренние компоненты, «переваривая» их ферментами лизосом. Это непрерывный процесс, поддерживающий баланс между синтезом и деградацией (распадом) и обеспечивающий необходимые условия для нормального клеточного роста, развития и смерти. Аутофагия (от греч. αυτος — «само» и φαγειν — «есть»: самопоедание) является клеточным механизмом утилизации избыточных или поврежденных белков, белковых комплексов и клеточных органелл, осуществляемый лизосомами той же клетки. Такая утилизация выполняет несколько важных функций, в том числе — получение питательных веществ при голодании, поддержку клеточного гомеостаза и клеточного иммунитета, осуществление апоптоза и т.п. (Daniel J. Klionsky, Fabio C. Abdalla, Hagai Abeliovich, Robert T. Abraham, Abraham Acevedo-Arozena, et. al.. (2012). Guidelines for the use and interpretation of assays for monitoring autophagy. Autophagy. 8, 445-544). Можно сказать, что аутофагия в узком смысле - это процесс, когда клетка подъедает что-то в самой себе с помощью ферментативного разложения этого чего-то. Главное действующее лицо в этом процессе - внутриклеточные органоиды лизосомы. Отличительной чертой аутофагической гибели клеток являются аутофагосомальные вакуоли, связанные с мембраной и характерные двойные мембраны. Аутофагия в широком смысле - на уровне любой биосистемыАутофагия - это широко распространённый среди живых систем на разных уровнях жизни процесс поглощения биологической системой какой-то своей части для поддержания собственной структуры и жизнедеятельности. Образно можно назвать это так: «самопожирание ради выживания». Клетки поедают (утилизируют) свои органоиды, фагоциты поедают клетки собственного организма, хрящевая ткань разрушается, поедается и замещается костной тканью, в процессе эмбрионального развития поедаются (разрушаются) временно существующие структуры. Важная функция аутофагии - обновление частей системы с целью поддержания ее стабильности как целого (гомеостаза). Стабильность биосистем достигается путем постоянного обновления компонентов через аутофагию. Непрерывная утилизация старых компонентов обновляет биосистему, а также позволяет пополнять энергетические запасы. К примеру, постаревшие за 3 месяца своего существования эритроциты разрушаются и уничтожаются, но их компоненты используются для создания и роста новых эритроцитов. Этот принцип используется в биосистемах на различных биологических уровнях. В эукариотической клетке отработавшие свой ресурс органеллы перевариваются лизосомами, уступая место новым. Отработавшие своё белки и матричные РНК разбираются на мономеры, которые затем собираются в новые белки и РНК. На уровне организма поврежденные клетки элиминируются посредством апоптоза или иммунной системой. В экосистемах отношения хищник–жертва не только поддерживают численность хищных видов, но и регулируют гомеостаз всей экосистемы, очищая её от слабых и больных животных и предохраняя виды от вырождения. Даже у прокариот известны «аутофагические» наклонности. Правда, они описаны не для отдельных клеток, а для прокариотических колоний под следующими названиями: бактериальный каннибализм, бактериальный альтруизм, аутолиз или запрограммированная клеточная смерть. Биологический механизм, запускающий аутофагию в этом случае, обнаружен у многих видов бактерий — это так называемая токсин-антитоксиновая система. Суть её состоит в том, что при голодании колония лизирует («переваривает») часть своих клеток для того чтобы остальные бактерии получили достаточно пищи для выживания. Таким образом колония переживает неМеханизм нетозахватку ресурсов или внешние неблагоприятные условия. При нехватке пищи часть бактерий в колонии выделяет в окружающую среду ядовитый токсин. При этом только некоторые из них способны производить молекулу антитоксина — белка, который обезвреживает токсин при его попадании в клетку. Такие клетки выживают и поглощают остальные, погибшие и лизированные под действием токсина. Это дает оставшимся в живых энергию, необходимую для споруляции и самосохранения. Вот такой получается бактериальный каннибализм. Аутолиз как частный случай аутофагии и часть процесса некроза Аутолиз - это процесс самопереваривания клеток, их растворение, расплавление за счёт действия собственных протеолитических ферментов без участия микроорганизмов. После смерти живого существа или его отдельного органа в его клетках начинается процесс самопереваривания. Ферменты выходят наружу из лизосом и/или митохондрий и начинают процессы расщепления органических веществ своей собственной клетки. Ферменты, содержащиеся в лизосомах, составляют единую функциональную группу и выполняют главным образом литическую функцию, т. е. функцию расщепления, переваривания. Они расщепляют все основные классы биологически активных соединений, в частности, нуклеиновые кислоты (РНК и ДНК) и полисахариды, а главное - белки. В митохондриях также имеются ферменты, которые разрушают главным образом продукты гликолиза, жирные кислоты и продукты распада аминокислот. Аутолиз бывает не только посмертным, но и прижизненным – в виде распада отдельных органоидов клетки, отдельных клеток или части ткани (органа) при сохранении основных жизненных функций целого организма. Аутолиз часто начинается ещё в период обратимых изменений, т. е. ещё до момента смерти клетки или организма. При жизни в нормальных условиях мембранные оболочки лизосом и митохондрий не дают ферментам выйти в цитоплазму и тем самым предохраняют клетку от их разрушающего действия. После смерти нарушается физико-химическое равновесие, в клетках происходит повреждение всех мембран и они становятся более проницаемыми. Выйдя из своих органоидов в цитоплазму, лизосомные и митохондриальные ферменты начинают аутолитический процесс в клетке. Мембрана лизосом вообще может повреждаться очень быстро при прекращении доступа кислорода. Но в некоторых органах (миокарде, почках, печени) лизосомы могут сохраняться неповрежденными до 10 часов после смерти. В целом, скорость аутолитических процессов пропорциональна степени активности и напряженности прижизненного метаболизма клеток. Чем активнее шёл обмен веществ в клетке - тем активнее и быстрее она будет подвергаться аутолизу. Аутолиз изменяет состояние и вид ядра клетки. Проявления аутолиза на светооптическом уровне под микроскопом:
Источник: Прижизненный некроз и посмертный аутолиз: проблема дифференциальной диагностики / Богомолова И.Н., Богомолов Д.В. // Избранные вопросы судебно-медицинской экспертизы. — Хабаровск, 2012 — №12. — С. 25-31 (https://www.forens-med.ru/book.php?id=1569). Органы в состоянии аутолиза дряблые, имеют тусклый вид, пропитаны кровяной плазмой. В пищевой промышленности аутолиз используется для придания нежной консистенции мясным и рыбным продуктам. В результате выдержки и засолки ткани мяса и рыбы за счёт аутолиза сами по себе размягчаются. 2. НекрозНекроз — это гибель части живого организма, необратимое отмирание его частей, тогда как целое — организм - остается живым. Напротив, термин "смерть" используется для обозначения прекращения жизнедеятельности всего организма в целом. Для клеточного некроза характерны расширенные митохондрии и клеточные органеллы из-за увеличения проницаемости клеточных мембран для небольших заряженных молекуhttps://ru.wikipedia.org/wiki/%D0%9D%D0%B5%D1%82%D0%BE%D0%B7л. Некроз (от греч. nekros — мёртвый) — это омертвение, гибель клеток и тканей в живом организме; при этом жизнедеятельность их полностью прекращается. Понятие "некроз" является видовым по отношению к более общему понятию "смерть". До недавнего времени некроз считался единственным вариантом смерти клетки в живом организме с хорошо изученными биохимическими, патофизиологическими, морфологическими и клиническими проявлениями. Однако в последние годы описан еще один вид смерти клетки в живом организме — это апоптоз, отличающийся от некроза тем, что он происходит по определенной генетической программе, имеющий особую биохи2+мическую и морфологическую сущность, а также клиническое значение. Некротические процессы происходят в организме постоянно, не только при патологии, но и в норме. Некротические процессы — закономерные проявления нормальной жизнедеятельности организма, так как для отправления любой физиологической функции требуются затраты материального субстрата (гибель клеток), которые постоянно восполняются путем физиологической регенерации. Кроме того, клетки постоянно подвергаются старению и естественной смерти с последующей их элиминацией. Таким образом, динамическое равновесие между процессами естественной смерти клеток и физиологической регенерацией обеспечивает постоянство клеточных и тканевых популяций в организме. Развитие очага некроза в ткани, как правило, сопровождается возникновением местной реакции — демаркационного острого воспаления, возникновение которого связывают с выделением некротизированной тканью провоспалительных субстанций. Предполагают генерацию погибающими клетками лейкотриенов — мощных медиаторов воспаления. Морфогенетические стадии некроза:
Наиболее достоверными из морфологических критериев необратимого повреждения клетки (некроза) являются нарушения в митохондриях, видимые под электронным микроскопом: разрушение внутренних мембран и отложения электронно-плотных депозитов, содержащих белки и соли кальция. На светооптическом уровне изменения в структуре клетки становятся видимыми лишь на стадии аутолиза. Поэтому, говоря о микроскопических признаках некроза, фактически говорят и о морфологических изменениях в стадии аутолиза, являющихся результатом действия гидролитических ферментов, прежде всего лизосомального происхождения. Кроме того, большинство органелл клетки (ядра, митохондрии, рибосомы и др.) также имеют свои собственные гидролитические ферменты, которые принимают активное участие в процессах аутолиза. Главные микроскопические признаки некроза. Заключаются в изменениях ядра и цитоплазмы клеток. Ядра последовательно подвергаются сморщиванию (кариопикноз), распаду на глыбки (кариорексис) и лизируются (кариолизис). Эти изменения ядер связаны с активацией гидролаз — рибонуклеаз и дезоксирибонуклеаз. В цитоплазме происходит денатурация и коагуляция белков, сменяемая обычо колликвацией. Коагуляция циоплазмы сменяется распадом её на глыбки (плазморексис) и лизисом (расплавлением) органелл (плазмолизис). При очаговых (фокальных) изменениях говорят о фокальном коагуляционном некрозе и фокальном колликвационном некрозе (баллонная дистрофия). Некроз развивается не только в паренхиматозных элементах тканей и органов, но и в их строме. При этом разрушаются как клетки стромы, так и нервные окончания и компоненты экстрацеллюлярного матрикса. Расщепление ретикулярных, коллагеновых и эластических волокон происходит с участием нейтральных протеаз (коллагеназ, эластазы), гликопротеидов — протеаз, липидов — липаз. При микроскопическом исследовании обнаруживаются распад, фрагментация и лизис ретикулярных, коллагеновых и эластических волокон (эластолизис), в некротизированной ткани нередко откладывается фибрин. Описанные изменения характерны для фибриноидного некроза. В жировой ткани некроз носит свои специфические черты в связи с накоплением в некротических массах жирных кислот и мыл, что ведёт к образованию липогранулём. Ультраструктурные признаки некроза отражают изменения органоидов клетки
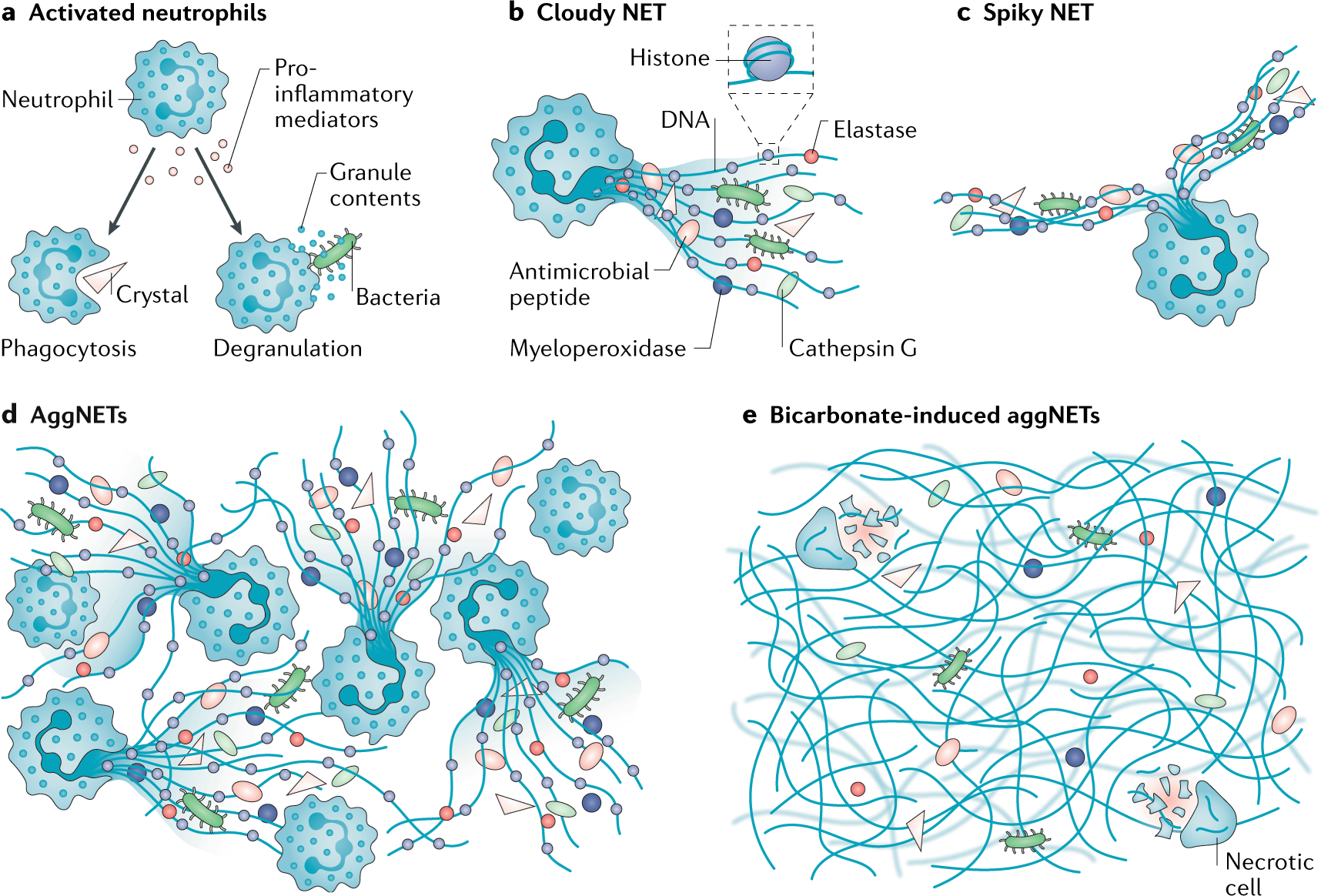
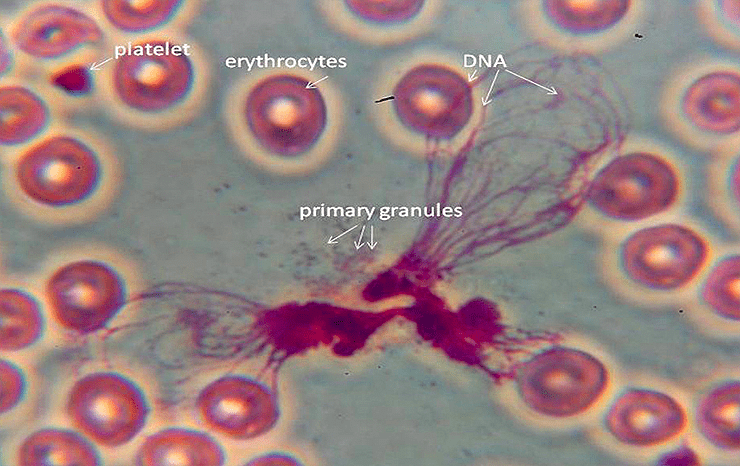
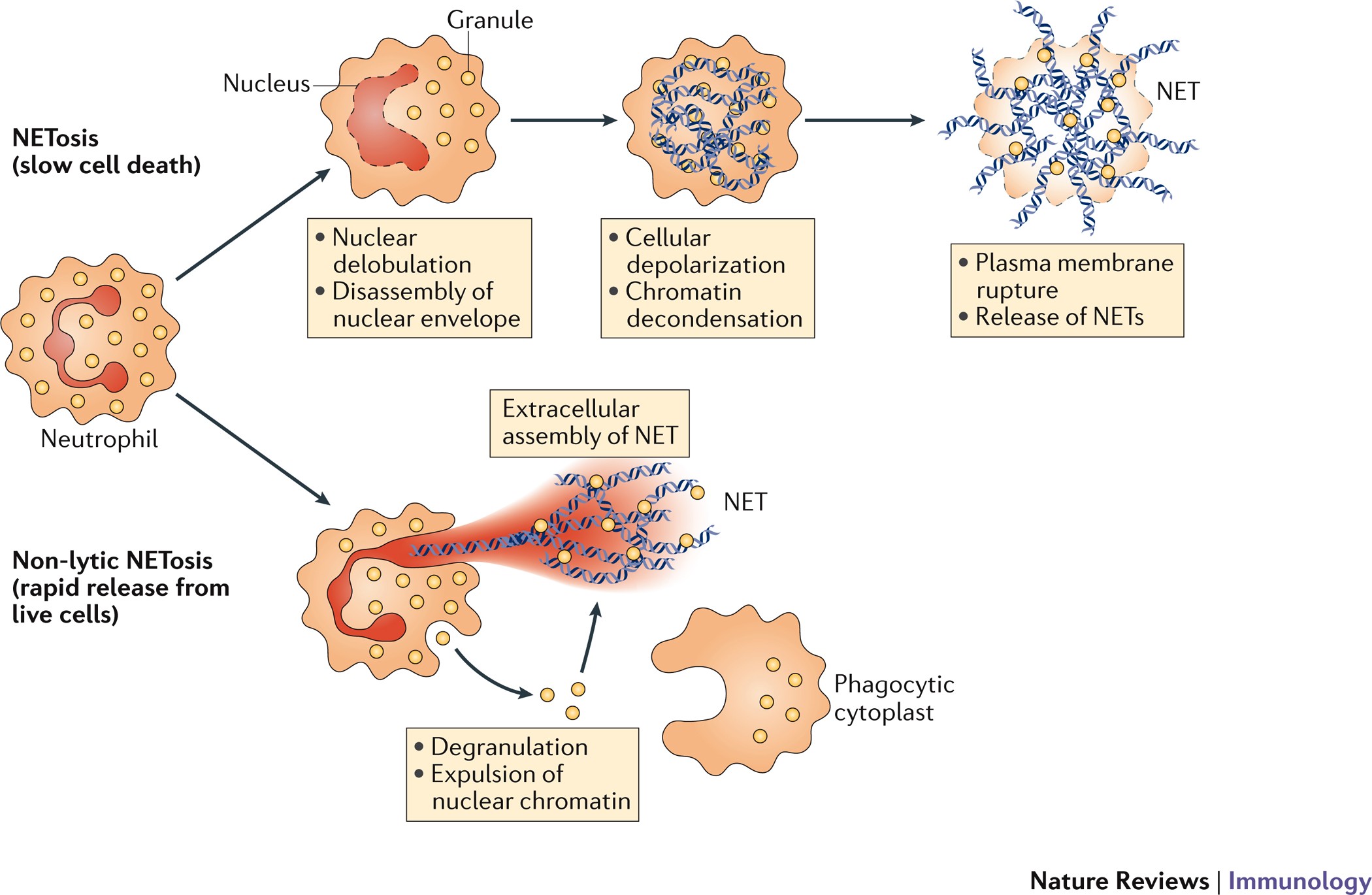
Конечный результат всех механизмов некроза — возникновение внутриклеточного хаоса. 3. Нетоз (НЕТоз) и этоз (ЭТоз)Этот вид "жертвенной" клеточной гибели характерен для лейкоцитов и лимфоцитов, но в наибольшей степени - для нейтрофилов. Рисунок 3 - Нейтрофилы выбрасывают во внешнюю среду самое дорогое - ДНК, превращая её во внеклеточные ловушки. Источник: https://media.springernature.com/full/springer-static/image/art%3A10.103... Нетоз не связан с фрагментацией ДНК или воздействием фосфатидилсерина (PS) на наружном слое клеточной мембраны, которые являются отличительными признаками апоптоза, поэтому мы не можем признать нетоз и этоз разновидностями апоптоза. Дополнительной особенностью, которая отличает нетоз от апоптоза и некроза, является то, что в его случае как ядерные, так и гранульные мембраны подвергаются фрагментации. Нейтрофилы (нейтрофильные гранулоциты) являются гетерогенной совокупностью пластичных гранулярных лейкоцитов с разнообразными функциями и фенотипическими характеристиками. Они составляют около 70% всех лейкоцитов человека. После выхода из красного костного мозга нейтрофилы человека живут от 4 часов до 5-6 суток и за это время "стареют". Часть старых нейтрофилов возвращается в красный костный мозг (ККМ), где они подвергаются апоптозу и фагоцитируются стромальными макрофагами. Другие по окончании своего срока жизни уничтожаются преимущественно макрофагами селезенки и печени. Нейтрофилы участвую в воспалительных реакциях, но кроме этого оказывают регуляторное влияние на другие клетки иммунной системы и развитие иммунного ответа. Рисунок 4 - Нейтрофил раскидывает свои внеклеточные ловушки среди других клеток крови. Истояник: https://biomolecula.ru/img/content/2218/03.netoz.png Нето́з (НЕТоз) (англ. NETosis от англ. Neutrophil extracellular traps - нейтрофильные внеклеточные ловушки) — вид программируемой клеточной гибели, происходящей у нейтрофилов, которая сопровождается выбрасыванием из погибающих нейтрофилов особых нитей, состоящих из молекул ДНК и связанных с ними белков. Благодаря этим нитям нейтрофилы убивают внеклеточных болезнетворных микробов (патогенов), минимизируя их вред для других клеток организма. Это явление действительно похоже на выбрасывание из клетки нейтрофила ловчих петель, или силков, предназначенных для поимки микробов. Ловчие петли (NETs) состоят из тяжей ДНК, связанной с глобулярными белковыми доменами, диаметром 15—17 нм и 25 нм соответственно. Тяжи сворачиваются в более толстые волокна диаметром 50 нм. Однако в некоторых условиях NETs могут формировать большие скопления, достигающие сотен нанометров в длину и ширину. Интересно то, что в данном случае ДНК выступает не в обычной для себя роли информационного носителя наследственной информации, а в роли эффекторной молекулы, которая сама "лично" отлавливает в межклеточной жидкости или крови микробов. Противомикробное действие NETs обусловлено тем, что они содержат белки с соответствующими свойствами — нейтрофилэластазу и катепсин G. Кроме того, входящие в NETs ядерные белки гистоны обладают высоким сродством к ДНК. Нити NETs обеспечивают повышенную локальную концентрацию молекул с антимикробными свойствами и обездвиживают клетки микробов. Кроме того, NETs образуют физический барьер, препятствующий распространению микробных клеток. NETs действуют не только на бактериальные клетки, но и на патогенные грибки, такие как дрожжи Candida albicans. В кровеносных сосудах на NETs могут застревать бактерии, циркулирующие по кровотоку. https://biomolecula.ru/img/content/2218/03.netoz.pngРисунок 5 - Суицидалльный и витальный виды нетоза. Источник: https://media.springernature.com/full/springer-static/image/art%3A10.103... Механизм нетозаПопав в "окружение" микробов нейтрофилы и другие лейкоциты выбрасывают из себя в окружающее пространство гранулы, состоящие из белков и хроматина, который, в свою очередь, состоит из ДНК. Во внеклеточном пространстве гранулы преобразуются в молекулярные нити (фибриллы), в которых запутываются чужеродные клетки. Нетоз не обязательно приводит к гибели нейтрофила. В случае витального нетоза молекулы ДНК вначале упаковываются в пузырьки-везикулы, которые отшнуровываются от клеточного ядра и затем выводятся наружу из клетки. Нейтрофил остаётся в живых, хотя и лишается своего ядра. Он даже может продолжать фагоцитировать бактерии и прочие частицы. Витальный нетоз запускается различными путями: липополисахаридами и другими антигенами бактериального происхождения, TLR4-активированными тромбоцитами или белками комплемента совместно с лигандами TLR2. При витальном нетозе происходит отшнуровывание пузырьков ядра (блеббинг), в результате чего в цитоплазме появляются везикулы, заполненные ДНК. Процесс их образования и выброса содержимого во внеклеточное пространство происходит очень быстро. Важно то, что нити ДНК выделяются наружу путём экзоцитоза без повреждения клеточной мембраны и нейтрофил при этом не погибает, хотя и остаётся без ДНК. Вопрос о том, можно ли считать живым такой нейтрофил, в котором отсутствует ДНК, является спорным, но по аналогии с эритроцитами его следует признать форменным элементом, лишённым ядра. Важно то, что после витального нетоза нейтрофилы могут продолжить свою полезную деятельность: фагоцитировать и убивать клетки микроорганизмов. Как обычно запускается и протекает нетоз? Нетоз запускается путём активации фермента NADPH-оксидазы, которая начинает производить активные формы кислорода. Под их действием запускается в работу другой фермент - протеин-аргининдеиминаза 4 (PAD4). PAD4 вызывает цитруллинирование белков гистонов в ядре нейтрофила. В результате этого хроматин теряет свою компактную укладку (декомпактизируется). Это означает, что в ядре заметный в виде глыбок хорошо окрашенный гетерохроматин превращается в слабоокрашенный эухроматин. В ядро входят ферменты нейтрофилов миелопероксидаза и эластаза. Они стимулируют дальнейшую декомпактизацию хроматина и в конце концов приводят к разрушению ядерной оболочки. Распутанный (деконденсированный) эухроматин выходит из разрушенного ядра в цитоплазму. Здесь к нему присоединяются дополнительные белки, формируя "ловчие петли" - внеклеточные нити нейтрофилов (англ. Neutrophil extracellular traps, NETs). Образование NETs регулируется липоксигеназным путём. При некоторых способах запуска нетоза (например, при контакте с бактериальной клеткой) нейтрофил-5-липоксигеназа взаимодействует с фосфолипидами, и продукты реакции угнетают образование NETs. В случае суицидального нетоза нейтрофил погибает. Суицидальный нетоз с выбрасыванием NETs приводит к гибели нейтрофилов. Этот механизм клеточного самоубийства отличается как от апоптоза, так и от некроза. В случае суицидального нетоза после образования в цитоплазме внеклеточных нитей ДНК у нейтрофила разрушается клеточная мембрана. В результате этого ловчие нити оказываются во внеклеточном пространстве. Но нейтрофил в качестве отдельной клетки перестаёт существовать. Суицидальный нетоз может быть запущен при активации у нейтрофила мембранных Toll-подобных рецепторов (TLRs), Fc-рецепторов и рецепторов комплемента различными лигандами, такими как антитела или форболмиристатацетат. Считается, что при активации этих рецепторов в конечном счёте происходит выход ионов кальция из цистерн эндоплазматической сети. Интересно, что процесс суицидального нетоза может занимать несколько часов, даже при воздействии высоких концентрации форболмиритатацетата, в то время как витальный нетоз происходит в течение всего нескольких минут. Ещё быстрее выбрасывают свои ловчие нити, состоящие из ДНК, эозинофилы - за несколько секунд. Но эозинофилы интересны не только этим, а также тем, что для своих внеклеточных ловушек они могут использовать не только ядерную, но и митохондриальную ДНК. Этоз эозинофилов предлагают называть ЭЭТозом. После завершения процесса "охоты на микробов" макрофаги убирают из внеклеточной среды ловчие нити NETs. Они фагоцитируют и разрушают эти нити. Нетоз может приносить не только пользу организму, но и вред. Так, выход гистонов во внеклеточную среду может запустить развитие аутоиммунных заболеваний, таких как системная красная волчанка. Также нетоз может участвовать в воспалительных заболеваниях. Итак, нетоз и его продукт NETs играют важную роль в патогенезе инфекционных, воспалительных и тромботических заболеваний. Другое применение ДНК "не по назначению"Особое применение своей ДНК нашли многоклеточные цианобактерии Anabaena, которые «реорганизовали» систему разделения коротких молекул ДНК (плазмид) в структуру цитоскелета, контролирующую форму этих клеток. В обычном случае система прокариот ParMR образует нити (филаменты) из актиноподобного белка, которые растаскивают малокопийные плазмиды к полюсам клетки. Однако у Anabaena эта система (переименованная в CorMR) не разделяет молекулы ДНК, а формирует из этой ДНК динамически нестабильные двухцепочечные нити на клеточной мембране, поддерживая форму клеток. Таким образом, ДНК формирует цитоскелет [Benjamin L. Springstein et al. , Repurposing of a DNA segregation machinery into a cytoskeletal system controlling cell shape. Science 392, eaea 6343 (2026). DOI:10.1126/science.aea6343, https://www.science.org/doi/10.1126/science.aea6343]. 4. АпоптозТермин"апоптоз", предложен в 1972 г. английскими учеными J.F.R. Кеrr, А.Н. Wyllie и A.R. Currie, состоит из двух греческих слов и означает в буквальном смысле "опадание лепестков цветов", а в применениии к клетке - особый тип гибели путем разделения её на части ("апоптозные тельца"), которые впоследствии фагоцитируются соседними клетками разного типа. Термин "программированная клеточная смерть" отражает функциональное назначение этого процесса, представляющего естественную часть функционирования многоклеточного организма, связанного с метаморфозом и развитием [ Hedgecock E.M., Salston J.E. 1983 , Oppenheim R.W. 1991 ]. Апопто́з (др.-греч. ἀπόπτωσις «опадание», от ἀπό- + πτῶσις «падение») — регулируемый процесс программируемой клеточной гибели. В результате клетка распадается на отдельные апоптотические тельца, ограниченные плазматической мембраной. Фрагменты погибшей клетки обычно очень быстро (в среднем за 90 минут фагоцитируются макрофагами либо соседними клетками, минуя развитие воспалительной реакции. Морфологически наблюдаемый процесс апоптоза продолжается 1—3 часа. Распространённость апоптоза. В организме среднестатистического взрослого человека в результате апоптоза погибает ежедневно порядка 50-70 миллиардов клеток (в два с лишним раза больше, чем количество нейронов в мозге). Для среднестатистического ребёнка в возрасте от 8 до 14 лет число клеток, погибших путём апоптоза, составляет порядка 20-30 миллиардов в день. Суммарная масса клеток, которые на протяжении одного года жизни подвергаются разрушению, эквивалентна массе тела человека. При этом восполнение утраченных клеток обеспечивается за счёт пролиферации — увеличения клеточной популяции путём деления. Образно можно сказать, что за год каждый человек "съедает" самого себя. Преднаначение апоптоза. Одной из основных функций апоптоза является уничтожение дефектных (повреждённых, мутантных, инфицированных) клеток. В многоклеточных организмах апоптоз к тому же задействован в процессах развития, дифференциации и морфогенеза. К примеру, утрата хвоста в процессе матаморфоза амфибий или атрофия у них гипохорды происходит благодаря массовому апоптозу целых клеточных популяций. Важна раль апоптоза в поддержании клеточного гомеостаза, в обеспечении развития и функционирования иммунной системы. Наприме, именно с помощью апоптоза осуществляют свои эффекторные функции цитотоксические лимфоциты: Т-киллеры и НК-клеки (натуральные киллеры). И те и другие клетки вводят внутрь клеток-мишеней сериновые протеазы (гранзимы), которые запускают в клетках-мишенях механизм апоптоза. Ктоме этого цитотоксические Т-лимфоциты способны запускать клеточную гибель своих мишеней путём активации у них мембранных рецепторов смерти. Результатом также является апоптоз. Апоптоз лежит в основе позитивной и негативной селекции (отбора) T- и B-лимфоцитов, в процессе самого формирования иммунитета. Апоптозу подвергаются иммунокомпетентные клетки, не прошедшие отбор. В гисто-гематических барьерах клетки, выполняющие барьерную функцию, запускают рецептор-зависимый апоптоз в тех эффекторных Т-лимфоцитах, которые пытаются проникнуть сквозь «барьерные» ткани. Главные ферменты-разрушители апоптоза. Каспазы - это цистеиновые протеазы, которые расщепляют в белках аминокислотные последовательности после остатка аспарагиновой кислоты. Можно назвать их цистеиновыми аспартазами, т.к. в активном центре у них цистеин, а мишенью-меткой для разрезания белков является аспартат. Апоптоз наблюдается у всех эукариотов, начиная от одноклеточных простейших и вплоть до высших организмов. В программируемой смерти прокариотов участвуют функциональные аналоги эукариотических белков апоптоза. В растительных организмах апоптоз столь же задействован в процессах морфогенеза и дифференциации, сколь и у животных. Программируемая клеточная смерть у растений обеспечивает ксилогенез и флоэмогенез, формообразование листьев, аэренхимогенез, образование корневого чехлика, опадание листьев и созревших плодов, прорастание пыльцевой трубки и т.д. Рисунок 6 - Фрагментированный лейкоцит в состоянии апоптоза. Источник изображения: https://edu.volgmed.ru/mod/book/tool/print/index.php?id=20308 В настоящее время установлены основные механизмы реализации апоптоза в эукариотических клетках, активно ведутся исследования регуляторов и активаторов апоптоза. Интерес учёных связан с тем, что механизмы апоптоза можно применить при лечении онкологических, аутоиммунных и нейродегенеративных заболеваний. Рисунок 7 - Схема апоптоза у млекопитающих. Источник изображения: https://edu.volgmed.ru/mod/book/tool/print/index.php?id=20308 Фазы апоптоза
1. Сигнальная фаза Запуск (инициация) апоптоза может происходить посредством внешних (внеклеточных) или внутриклеточных факторов. Например, в результате гипоксии, гипероксии, субнекротического поражения химическими или физическими агентами, перекрёстного связывания соответствующих рецепторов, нарушения сигналов клеточного цикла, удаления факторов роста и метаболизма и т. д. Несмотря на разнообразие инициирующих факторов, выделяются два основных пути передачи сигнала апоптоза: рецептор-зависимый (внешний) сигнальный путь с участием рецепторов гибели клетки и митохондриальный (собственный) путь Внешний рецептор-зависимый сигнальный путь Процесс апоптоза часто (например, у млекопитающих) начинается с присоединения специфических внеклеточных лигандов к специальным мембранным рецепторам клеточной гибели, выставленным (экспрессированным) на поверхности мембраны. Рецепторы, воспринимающие сигнал апоптоза, относятся к суперсемейству TNF-рецепторов (англ. tumor necrosis factor receptor или кратко TNFR — «рецептор фактора некроза опухолей»). Наиболее изученными рецепторами смерти, для которых описана и определена роль в апоптозе, являются CD95 (также известный как Fas или APO-1) и TNFR1 (также называемый p55 или CD120a). К дополнительным относятся CARI, DR3 (англ. death receptor 3 — «рецептор смерти 3»), DR4 и DR5. Все рецепторы смерти представляют собой трансмембранные белки, характеризующиеся наличием общей последовательности из 80 аминокислот в цитоплазматическом домене. Данная последовательность называется доменом смерти (англ. death domain или кратко DD) и является необходимой для преобразования (трансдукции) сигнала апоптоза. Внеклеточные участки рецепторов смерти взаимодействуют с тримерами лигандов (CD95L, TNF, Apo3L, Apo2L и т. п.). Тримеры лигандов в результате взаимодействия тримеризуют рецепторы смерти (то есть «сшивают» 3 молекулы рецептора). Активированный таким образом рецептор взаимодействует с соответствующим внутриклеточным адаптером (или адаптерами). Для рецептора CD95 (Fas/APO-1) адаптером является FADD (от англ. Fas-associated DD-protein — «белок, взаимодействующий с доменом смерти Fas-рецептора»). Для рецепторов TNFR1 и DR3 адаптером является TRADD (от англ. TNFR1-associated DD-protein — «белок, взаимодействующий с доменом смерти TNFR1-рецептора»). Адаптер, ассоциированный с рецептором смерти, вступает во взаимодействие с эффекторами — пока ещё неактивными предшественниками протеаз из семейства инициирующих каспаз — с прокаспазами. В результате цепочки взаимодействия «лиганд-рецептор-адаптер-эффектор» формируются агрегаты, в которых происходит активация каспаз. Данные агрегаты именуются апоптосомами, апоптозными шаперонами или сигнальными комплексами, индуцирующими смерть (от англ. DISC — death-inducing signaling complex — «сигнальный комплекс, индуцирующий смерть»). Примером апоптосомы может служить комплекс FasL-Fas-FADD-прокаспаза-8, в котором активируется каспаза-8. Рецепторы смерти, адаптеры и эффекторы взаимодействуют между собой сходными по структуре доменами: DD, DED, CARD. DD (от англ. death domain — «домен смерти») участвует во взаимодействии рецептора Fas с адаптером FADD и во взаимодействии рецепторов TNFR1 или DR3 с адаптером TRADD. Посредством домена DED (от англ. death-effector domain — «домен эффектора смерти») осуществляется взаимодействие адаптера FADD с прокаспазами −8 и −10. Домен CARD (от англ. caspase activation and recruitment domain — «домен активации и рекрутирования каспазы») участвует во взаимодействии адаптера RAIDD с прокаспазой-2. Посредством рецепторов смерти могут быть активированы три инициирующие каспазы: −2; −8 и −10. Активированные инициирующие каспазы далее участвуют в активации эффекторных каспаз. Митохондриальный сигнальный путь Большинство форм апоптоза у позвоночных реализуется не по рецепторному, а по митохондриальному пути. Митохондриальный сигнальный путь апоптоза реализуется в результате выхода апоптогенных белков из межмембранного пространства митохондрий в цитоплазму клетки. Высвобождение апоптогенных белков, предположительно, может осуществляться двумя путями: за счёт разрыва митохондриальной мембраны или же путём открытия высокопроницаемых каналов на внешней мембране митохондрий. Ключевым событием митохондриального пути апоптоза является повышение проницаемости наружной мембраны митохондрий (англ. mitochondrial outer membrane permeabilization, MOMP). Существенную роль в повышении MOMP играют апоптотические Bcl-2-белки — Bax и Bak. Они встраиваются в наружную мембрану митохондрий и олигомеризуются. При этом, вероятно, нарушается целостность внешней мембраны митохондрий. При повышении MOMP из межмембранного пространства митохондрий в цитозоль высвобождаются растворимые белки, участвующие в апоптозе: цитохром c — белок с молекулярной массой 15 кДа; прокаспазы −2, −3 и −9; белок AIF (англ. apoptosis inducing factor — «фактор, индуцирующий апоптоз») — флавопротеин с молекулярной массой 57 кДа. Разрыв внешней мембраны митохондрий объясняется увеличением объёма митохондриального матрикса. Данный процесс связывают с раскрытием пор митохондриальной мембраны, приводящим к снижению мембранного потенциала и высокоамплитудному набуханию митохондрий вследствие осмотического дисбаланса. Поры диаметром 2,6—2,9 нм способны пропускать низкомолекулярные вещества массой до 1,5 кДа. Раскрытие пор стимулируют следующие факторы: неорганический фосфат; каспазы; SH-реагенты; истощение клеток восстановленным глутатионом; образование активных форм кислорода; разобщение окислительного фосфорилирования протонофорными соединениями; увеличение содержания Ca2+ в цитоплазме; воздействие церамида; истощение митохондриального пула АТФ и др. Цитохром c в цитоплазме клетки участвует в формировании апоптосомы вместе с белком APAF-1 (от англ. Apoptosis Protease Activating Factor-1 — «активирующий фактор апоптотической протеазы-1»). Предварительно APAF-1 претерпевает конформационные изменения в результате реакции, протекающей с затратой энергии АТФ. Предполагается, что трансформированный APAF-1 приобретает способность связывать цитохром c. К тому же открывается доступ CARD-домена APAF-1 для прокаспазы-9. В итоге происходит олигомеризация 7 субъединиц трансформированного белка APAF-1 с участием цитохрома c и прокаспазы-9. Так образуется апоптосома, активирующая каспазу-9. Зрелая каспаза-9 связывает и активирует прокаспазу-3 с образованием эффекторной каспазы-3.[22] Высвобождающийся из межмембранного пространства митохондрий флавопротеин AIF является эффектором апоптоза, действующим независимо от каспаз. Другие пути индукции апоптоза Реализация апоптоза может происходить в результате комбинированного действия двух основных сигнальных путей: рецептор-зависимого и митохондриального. Но помимо этого существует ряд менее распространённых механизмов инициации апоптоза. Например, за счёт активации прокаспазы-12, локализованной в эндоплазматическом ретикулуме. Высвобождение и активация прокаспазы-12 при этом обусловлены нарушениями внутриклеточного гомеостаза ионов кальция (Ca2+). Активация апоптоза также может быть связана с нарушением адгезии (прилипания) клеток. Т-киллеры. В качестве ещё одного фактора индукции апоптоза рассматривается атака инфицированных клеток цитотоксическими Т-лимфоцитами, которые, помимо активации Fas-рецептора, способны секретировать перфорин вблизи мембраны заражённой клетки. Перфорин, полимеризуясь, образует трансмембранные каналы, через которые внутрь клетки поступают лимфотоксин-альфа и смесь сериновых протеаз (гранзимов). Далее гранзим B активирует каспазу-3 и запускается каспазный каскад. Возможна инициация клеточной смерти при высвобождении лизосомальных протеаз — катепсинов. К примеру, каспаза-8 вызывает выход из лизосом активного катепсина B, который затем расщепляет регуляторный белок Bid. В результате образуется активный белок t-Bid, активирующий в свою очередь проапоптозный белок Bax. 2. Эффекторная фазаВ течение эффекторной фазы различные инициирующие пути сходятся (конвертируются) в один общий путь апоптоза (или несколько путей). Как правило, происходит активация каскада белков-эффекторов и регулирующих их белков-модуляторов. Основными эффекторами апоптоза являются каспазы. В процессе активации они запускают каспазный каскад: сложно переплетённые цепочки взаимодействий инициирующих и эффекторных каспаз. Каспазный каскад Ферменты-разрушители - каспазы. Каспазы образуются за счёт активации прокаспаз (молекулярная масса 32—56 кДа), в составе которых выделяют 3 домена: регуляторный N-концевой домен (продомен), большую (17—21 кДа) и малую (10—13 кДа) субъединицы. Активация происходит путём протеолитического процессинга: все три домена расщепляются, отделяется продомен, а оставшиеся большая и малая субъединицы ассоциируются, образуя гетеродимер. Два гетеродимера в дальнейшем формируют тетрамер — полноценную каспазу с двумя каталитическими участками. Каспазы обнаружены в большинстве живых организмов. У млекопитающих идентифицировано 13 каспаз. Часть из них в апоптозе не участвует (−1, −4, −5, −11, −13). Остальные каспазы, которые участвуют в апоптозе, разделяют на инициаторные (−2, −8, −9, −10, −12) и эффекторные (исполнительные) (−3, −6, −7). Инициаторные каспазы активируют эффекторные каспазы, которые в свою очередь провоцируют и непосредственно участвуют в трансформации клетки. В итоге морфологические и биохимические изменения приводят к гибели клетки по типу апоптоза. Одна из основных функций эффекторных каспаз заключается в прямом и опосредованном разрушении клеточных структур. Гидролизу подвергаются белки ядерной ламины, разрушается цитоскелет, расщепляются белки, регулирующие клеточную адгезию. Другой важной функцией эффекторных каспаз является инактивация белков, блокирующих апоптоз. В частности расщепляется ингибитор DFF (англ. DNA fragmentation factor — «фактор фрагментации ДНК»), препятствующий активации апоптозной ДНКазы CAD (англ. caspase-activated DNase — «ДНКаза, активируемая каспазами»). Разрушению подвергаются и антиапоптозные белки семейства Bcl-2. Наконец, в результате действия эффекторных каспаз происходит диссоциация регуляторных и эффекторных доменов, участвующих в репарации ДНК, мРНК-сплайсинга и ДНК-репликации. Помимо каспаз существуют и другие эффекторы апоптоза. Например, флавопротеин AIF, высвобождающийся из межмембранного пространства митохондрий, действует по независимому от каспаз пути. Попадая в клеточное ядро, AIF вызывает конденсацию хроматина и активирует эндонуклеазы, которые участвуют во фрагментации ДНК. На основании экспериментальных данных установлено, что апоптоз, протекающий в присутствии AIF, не предотвращается ингибитором каспаз (Z-VAD-fmk). В качестве эффекторов апоптоза также рассматриваются кальпаины — представители семейства цитозольных Ca2+-активируемых цистеиновых протеаз. 3. Деградационная фазаИтогом программируемой клеточной гибели вне зависимости от изначального инициирующего воздействия является деградация клетки путём фрагментации на отдельные апоптотические тельца, ограниченные плазматической мембраной. Фрагменты погибшей клетки обычно очень быстро (в среднем за 90 минут) фагоцитируются макрофагами либо соседними клетками, минуя развитие воспалительной реакции. Дело в том, что на внешней стороне мембраны апоптотических телец появляется вещество фоспатидилсерин (PS), который является маркером апоптоза. Фагоциты опознают фоспатидилсерин и поглощают меченые им фрагменты распавшихся клеток. С морфологической точки зрения деградацию погибающей от апоптоза клетки можно разделить на три последовательных фазы: высвобождение, блеббинг и конденсация. Деградация большинства клеток начинается с высвобождения прикреплений внеклеточного матрикса и реорганизации фокальной адгезии. Внутри погибающей клетки деполимеризуются микротрубочки цитоскелета. Внутриклеточные актиновые микрофиламенты реорганизуются в связанные с мембраной периферийные (кортикальные) кольцевые пучки. В итоге клетка приобретает округлую форму. Следующая за высвобождением стадия блеббинга характеризуется сокращением периферийных актиновых колец. В результате сокращений клеточная мембрана образует вздутия, клетка как бы «пузырится». Процесс блеббинга энергозависим и требует большого количества АТФ. Фаза блеббинга в нормальных условиях завершается примерно через 1 час. В итоге клетка фрагментируется на маленькие апоптотические тела, либо целиком конденсируется, округляясь и уменьшаясь в размерах. На молекулярном уровне одним из последствий апоптоза является фрагментация ДНК с участием нуклеаз. Изначально образуются крупные фрагменты с 30000-700000 пар оснований, которые в дальнейшем расщепляются в межнуклеосомной области на отрезки по 180-200 пар оснований или кратные этим величинам. Фрагментация ДНК является характерным, но не обязательным признаком апоптоза, так как существуют наблюдения, в ходе которых процесс фрагментации ядра (кариорексис) протекал без сопутствующей фрагментации ДНК. Ещё одним существенным последствием апоптоза является экспрессия на внешней стороне плазматической мембраны специфических молекулярных маркеров, распознаваемых фагоцитирующими клетками: тромбоспондина; фосфатидилсерина и других фосфолипидов, содержащих фосфосерин. Маркеры, участвующие в фагоцитозе погибающей клетки, можно условно разделить на три группы: «ешь меня», «найди меня» и «не ешь меня». Критический сигнал «ешь меня» возникает при появлении на внешней стороне мембраны (экстернализации) фосфатидилсерина. В обычных условиях он локализуется на внутреннем слое плазматической мембраны. Данное состояние обеспечивается АТФ-зависимой флиппазой, которая перемещает фосфолипид с внешнего на внутренний слой. Во время апоптоза фосфатидилсерин, наоборот, перемещается на внешний слой плазматической мембраны при участии каспаз. Там он ассоциируется с такими белками, как аннексин I и MFG-E8, которые участвуют во взаимодействии с рецепторами фагоцитирующих клеток. В процессе апоптоза также снижается интенсивность сигналов «не ешь меня» (CD31), которые в норме присутствуют у фагоцитов и у большинства здоровых клеток. Третья группа сигналов «найди меня» (лизофосфатидилхолин) продуцируется гибнущей клеткой для привлечения фагоцитов: эффекторные каспазы активируют фосфолипазу A, которая участвует в образовании лизофосфатидилхолина. Различия между некрозом и апоптозомРисунок 8 - Сравнение некроза и апоптоза. Источник изображения: https://ppt-online.org/202117 Сравнение апоптоза и некроза Перейти Отличия апоптоза от некроза связаны с различиями в их распространенности, генетическими, биохимическими, морфологическими и клиническими проявлениями. Существенным отличием является то, что некроз может захватывать территорию тела, начиная от части клетки до целого органа. Апоптоз же распространяется всегда только на отдельные клетки или их группы. Апоптоз возникает в клетках при определенных генетических перестройках. При апоптозе усиливается экспрессия (проявление) генов, контролирующих пролиферацию (размножение) и дифференцировку клеток. Это гены из группы клеточных онкогенов (c-fos, c-myc, c-bcl-2) и антионкогенов (р53). Активация клеточных онкогенов должна вести к усилению пролиферации клеток, однако при одновременной активации антионкогена р53 происходит не пролиферация, а апоптоз. Такие взаимоотношения генов демонстрируют возможность координации процессов пролиферации и гибели клеток, заложенной в генетическом аппарате клеток. В отличие от некроза разрушение ядра при апоптозе происходит с участием специальных Са2+, Mg2+-зaвисимыix эндонуклеаз, расщепляющих молекулы ДНК в участках между нуклеосомами. Это приводит к формированию однотипных по размерам фрагментов ДНК. Масса этих фрагментов кратна массе одной нуклеосомы, а каждый фрагмент содержит от одной до нескольких нуклеосом. Своеобразное расщепление ДНК при апоптозе имеет и свое морфологическое выражение в виде особой структуры хроматина. В цитоплазме клетки, подвергшейся апоптозу, никогда не наблюдается активации гидролитических ферментов, как это бывает при некрозе. Напротив, все органеллы долгое время остаются сохранными и подвергаются конденсации (сжатию), что связывают с процессами сшивания белковых молекул трансглютаминазами, а также обезвоживания за счет действия особых селективных ферментных транспортных систем, регулирующих обмен ионов калия, натрия, хлора и воды. При световой микроскопии клетки в состоянии апоптоза и их фрагменты (апоптозные тельца) отличаются небольшими размерами, сравнимыми с размерами лимфоцитов, с высоким ядерно-цитоплазматическим соотношением, округлыми контурами и конденсированными хроматином и цитоплазмой. Существенным отличием является также отсутствие воспалительной реакции на апоптоз. В отличие от некроза при апоптозе имеется сохранность и интегративность органелл. Митохондрии не набухают, в них не происходит разрыва внутренней мембраны. Характерными для апоптоза являются такие ультраструктурные изменения, как агрегация рибосом в полукристаллоидные структуры, появление пучков микрофиламентов под цитолеммой, расположенных параллельно мембране. Почти всегда наблюдается кратковременная дилатация агранулярной эндоплазматической сети с формированием пузырей, наполненных жидкостью, которые выводятся из клетки. Наиболее яркое отличие апоптоза от некроза связано с изменениями ядерного хроматина, который конденсируется под кариолеммой в виде полусфер и глыбок. В ядре обнаруживаются осмиофильные тельца, сформированные транскрипционными комплексами, поступающими из ядрышек. Ядро меняет свою форму, становится изрезанным, фрагментирустся, ядерные поры концентрируются только в участках, где отсутствует маргинации хроматина. Клетка в состоянии апоптоза становится привлекательным объектом фагоцитоза для соседних паренхиматозных и стромальных клеток в прежде всего для макрофагов. Фагоцитоз происходит настолько быстро, что в условиях in vivo апоптозные клетки сохраняются всего в течение нескольких минут, что затрудняет их наблюдение. Рисунок 9 - Упрощённая схема апоптоза. Источник изображения: https://ppt-online.org/202117 Примеры естественного апоптоза: возрастная атрофия тимуса, возрастная инволюция ткани эндометрия и предстательной железы, молочных желёз после прекращения лактации. Классическим примером может также служить апоптоз В- и Т-лимфоцитов после прекращения стимулирующего действия на них соответствующих цитокинов при завершении иммунных реакций. Видео: Апоптоз Видео: Реальный апоптоз Видео: Апоптоз
Источники: https://edu.volgmed.ru/mod/book/tool/print/index.php?id=20308 https://biomolecula.ru/articles/autofagiia-protofagiia-i-ostalnye#source-1 http://www.tiensmed.ru/news/apoptoz-c9t.html http://medbiol.ru/medbiol/apon/00029da8.htm http://www.rsmu.ru/fileadmin/rsmu/img/about_rsmu/disser/14/d_vikulina_as... https://ru.wikipedia.org/wiki/%D0%9D%D0%B5%D1%82%D0%BE%D0%B7 Wartha F, Henriques-Normark B. ETosis: a novel cell death pathway. Sci Signal. 2008 May 27;1(21):pe25. doi: 10.1126/stke.121pe25. PMID: 18506034. Wang LY, Liu XJ, Li QQ, Zhu Y, Ren HL, Song JN, Zeng J, Mei J, Tian HX, Rong DC, Zhang SH. The romantic history of signaling pathway discovery in cell death: an updated review. Mol Cell Biochem. 2023 Oct 18. doi: 10.1007/s11010-023-04873-2. Epub ahead of print. PMID: 37851176.
Ваша оценка:
|
Пользователей онлайн: 0. Всего гостей: 0 Поисковики: нет. 
Приветствую вас на своем сайте, здесь вы можете найти много полезной информации (или что-то типа того) САЗОНОВ Вячеслав Фёдорович доцент кафедры биологии Рязанского государственного университета имени С.А. Есенина, кандидат биологических наук. Преподаватель вуза с 1978 года... РекламаПоискПритча наудачу:На сайте введена регистрация через социальные сети, если вы хотите оставлять комментарии без потверждения, пожалуйста, воспользуйтесь именно этим типом аутентификации. Если у вас уже есть аккаунт на сайте, вы можете привязать его к любой социальной сети? зайдя в настройки вашего аккаунта("Мои учётные данные") ниже и воспользовавшись вкладкой "Подключение к социальным сетям". После того, как вы зайдёте при помощи аккаунта в социальной сети, ваши возможности на сайте возрастут. Поддержка сайтаДанный сайт "Кинезиолог" (kineziolog.su) - продукт чисто человеческой "мозговой работы" (термин предложен автором сайта по аналогии с термином "ручная работа" или «handmade»): "мозговая работа" = «brainmade» © 2026 Сазонов В.Ф. © 2026 kineziolog.su). Это означает, что данный сайт создан и поддерживается за счёт чисто человеческой умственной деятельности, без использования нейросетей и/или искусственного интеллекта. Вы можете поддержать сайт своим добрым словом, лайком и/или полноценной ссылкой на его материалы! Это очень поможет. В ответ на вашу поддержку IT-специалисты, следящие за сайтом день и ночь, хотя бы удовлетворённо улыбнутся. ) -26-07-06-18-13 |









 ВКонтакте
ВКонтакте Facebook
Facebook Twitter
Twitter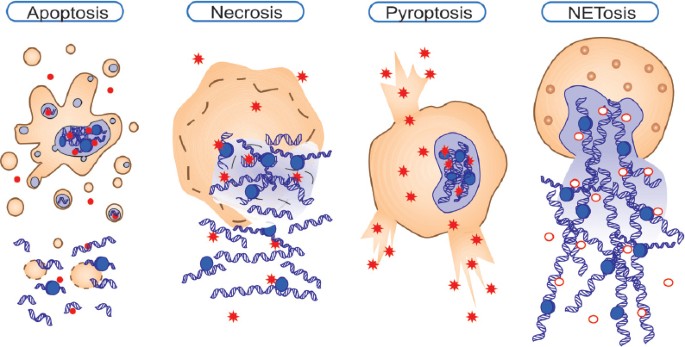{kind=link}
{kind=link}
{kind=link}
{kind=link}