






Гормональная регуляцияКраткое описание: Учебный материал по биохимии, молекулярной биологии и биорегуляции: Гормональная регуляция. МОДУЛЬ 11: ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНА ВЕЩЕСТВ И ФУНКЦИЙ ОРГАНИЗМА
Модульная единица 1 РОЛЬ ГОРМОНОВ В РЕГУЛЯЦИИ МЕТАБОЛИЗМА. РЕГУЛЯЦИЯ ОБМЕНА УГЛЕВОДОВ, ЛИПИДОВ, АМИНОКИСЛОТ ПРИ НОРМАЛЬНОМ РИТМЕ ПИТАНИЯ Цели изучения Уметь: 1. Применять знания о молекулярных механизмах регуляции обмена веществ и функций организма для понимания биохимических основ гомеостаза и адаптации. 2. Использовать знания о механизмах действия гормонов (инсулина и контринсулярных гормонов: глюкагона, кортизола, адреналина, соматотропина, йодтиронинов) для характеристики изменений энергетического обмена при смене периодов пищеварения и постабсорбтивного состояния. 3. Анализировать изменения метаболизма при гипо- и гиперпродукции кортизола и гормона роста, болезнь и синдром Иценко-Кушинга (акромегалия), а также при гипер- и гипофункции щитовидной железы (диффузный токсический зоб, эндемический зоб). Знать: 1. Современную номенклатуру и классификацию гормонов. 2. Основные этапы передачи гормональных сигналов в клетку. 3. Этапы синтеза и секреции инсулина и основных контринсулярных гормонов. 4. Механизмы поддержания в крови концентрации основных энергоноси- телей при нормальном ритме питания. Тема 11.1. РОЛЬ ГОМОНОВ В РЕГУЛЯЦИИ МЕТАБОЛИЗМА 1. Для нормального функционирования многоклеточного организма необходима взаимосвязь между отдельными клетками, тканями и органами. Эту взаимосвязь осуществляют: • нервная система (центральная и периферическая) через нервные импульсы и нейромедиаторы; • эндокринная система через эндокринные железы и гормоны, которые синтезируются специализированными клетками этих желез, выделяются в кровь и транспортируются к различным органам и тканям; • паракринная и аутокринная системы посредством различных соединений, которые секретируются в межклеточное пространство и взаимодействуют с рецепторами либо близлежащих клеток, либо той же клетки (простагландины, гормоны желудочно-кишечного тракта, гистамин и др.); • иммунная система через специфические белки (цитокины, антитела). 2. Эндокринная система обеспечивает регуляцию и интеграцию метаболизма в разных тканях в ответ на изменения условий внешней и внутренней среды. Гормоны функционируют как химические посредники, переносящие информацию об этих изменениях в различные органы и ткани. Ответная реакция клетки на действие гормона определяется как химическим строением гормона, так и типом клетки, на которую направлено его действие. Гормоны присутствуют в крови в очень низкой концентрации, и их действие обычно кратковременно. Это обусловлено, во-первых, регуляцией их синтеза и секреции и, во-вторых, высокой скоростью инактивации циркулирующих гормонов. Основные связи между нервной и эндокринной системами регуляции осуществляются с помощью специальных отделов мозга - гипоталамуса и гипофиза. В системе нейрогуморальной регуляции существует своя иерархия, вершиной которой является ЦНС и строгая последовательность протекания процессов. 3. Иерархия регуляторных систем. Системы регуляции обмена веществ и функций организма образуют три иерархических уровня (рис. 11.1). Первый уровень - центральная нервная система. Нервные клетки получают сигналы, поступающие из внешней и внутренней среды, преобразуют их в форму нервного импульса, который в синапсе вызывает освобождение медиатора. Медиаторы вызывают изменения метаболизма в эффекторных клетках через внутриклеточные механизмы регуляции. Второй уровень - эндокринная система - включает гипоталамус, гипофиз, периферические эндокринные железы, а также специализированные клетки некоторых органов и тканей (ЖКТ, адипоциты), синтезирующие гормоны и высвобождающие их в кровь при действии соответствующего стимула. Третий уровень - внутриклеточный - составляют изменения метаболизма в пределах клетки или отдельного метаболического пути, происходящие в результате: • изменения активности ферментов путем активации или ингибирования; • изменения количества ферментов по механизму индукции или репрессии синтеза белков или изменения скорости их деградации; • изменения скорости транспорта веществ через мембраны клеток. Синтез и секреция гормонов стимулируется внешними и внутренними сигналами, поступающими в ЦНС. Эти сигналы по нервным связям поступают в гипоталамус, где стимулируют синтез пептидных гормонов (так называемых рилизинг-гормонов) - либеринов и статинов. Либерины и статины транспортируются в переднюю долю гипофиза, где стимулируют или тормозят синтез тропных гормонов. Тропные гормоны гипофиза стимулируют синтез и секрецию гормонов периферических эндокринных желез, которые поступают в общий кровоток. Некоторые гипоталамические гормоны сохраняются в задней доле гипофиза, откуда секретируются в кровь (вазопрессин, окситоцин).
ТЕМА 11.2. МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНЫХ СИГНАЛОВ В КЛЕТКИ Биологическое действие гормонов проявляется через их взаимодействие с клетками, имеющими рецепторы к данному гормону (клетками-мишенями). Для проявления биологической активности связывание гормона с рецептором должно приводить к образованию химического сигнала внутри клетки, который вызывает специфический биологический ответ, например, изменение скорости синтеза ферментов и других белков или изменение их активности (см. модуль 4). Мишенью для гормона могут служить клетки одной или нескольких тканей. Воздействуя на клетку-мишень, гормон вызывает специфическую ответную реакцию, проявление которой зависит от того, какие метаболические пути активируются или тормозятся в этой клетке. Например, щитовидная железа - специфическая мишень для тиреотропина, под действием которого увеличивается количество ацинарных клеток щитовидной железы, повышается скорость биосинтеза тиреоидных гормонов. Глюкагон, воздействуя на адипоциты, активирует липолиз, в печени стимулирует мобилизацию гликогена и глюконеогенез. • Рецепторы гормонов могут быть расположены или в плазматической мембране или внутри клетки (в цитозоле или ядре). • По механизму действия гормоны можно разделить на две группы: - к первой группе относятся гормоны, взаимодействующие с мембранными рецепторами (пептидные гормоны, адреналин, а также гормоны местного действия - цитокины, эйкозаноиды); - вторая группа включает гормоны, взаимодействующие с внутриклеточными рецепторами - стероидные гормоны, тироксин (см. модуль 4). Связывание гормона (первичного мессенджера) с рецептором приводит к изменению конформации рецептора. Эти изменения улавливается другими макромолекулами, т.е. связывание гормона с рецептором приводит к сопряжению одних молекул с другими (трансдукция сигнала). Таким образом, генерируется сигнал, который регулирует клеточный ответ. В зависимости от способа передачи гормонального сигнала скорость реакций метаболизма в клетках меняется: • в результате изменения активности ферментов; • в результате изменения количества ферментов (рис. 11.2).
ТЕМА 11.3. СТРОЕНИЕ И БИОСИНТЕЗ ГОРМОНОВ 1. Пептидные гормоны синтезируются, как и другие белки, в процессе трансляции из аминокислот. Некоторые пептидные гормоны - это короткие пептиды; например, гормон гипоталамуса тиреотропин - либерин - трипептид. Большинство гормонов передней доли гипофиза - гликопротеины. Некоторые пептидные гормоны являются продуктами общего гена (рис. 11.3). Большинство полипептидных гормонов синтезируется в виде неактивных предшественников - препрогормонов. Образование активных гормонов происходит путем частичного протеолиза. 2. Инсулин - полипептид, состоящий из двух полипептидных цепей. Цепь А содержит 21 аминокислотный остаток, цепь В - 30 аминокислотных остатков. Обе цепи соединены между собой двумя дисульфидными мостиками. Молекула инсулина содержит также внутримолекулярный дисульфидный мостик в А-цепи. Биосинтез инсулина начинается с образования неактивных предшественников, препроинсулина и проинсулина, которые в результате последовательного протеолиза превращаются в активный гормон. Биосинтез препроинсулина начинается с образования сигнального пептида на полирибосомах, связанных с эндоплазматическим ретикулумом. Сигнальный
А - ПОМК (проопиомеланокортин) синтезируется в передней и промежуточной долях гипофиза и в некоторых других тканях ( кишечнике, плаценте). Полипептидная цепь состоит из 265 аминокислотных остатков; Б - после отщепления N-концевого сигнального пептида полипептидная цепь расщепляется на два фрагмента: АКТГ (39 а.к.) и β-липотропин (42-134 а.к.); В, Г, Д - при дальнейшем протеолизе происходит образование α- и β-МСГ (Меланоцитстимулирующего гормона) и эндорфинов. КППДГ - кортикотропиноподобный гормон промежуточной доли гипофиза. Процессинг ПОМК в передней и промежуточной долях гипофиза протекает по-разному, с образованием разного набора пептидов пептид проникает в просвет эндоплазматического ретикулума и направляет в ЭР растущую полипептидную цепь. После окончания синтеза препроинсулина сигнальный пептид отщепляется (рис. 11.4). Проинсулин (86 аминокислотных остатков) поступает в аппарат Гольджи, где под действием специфических протеаз расщепляется в нескольких участках с образованием инсулина (51 аминокислотный остаток) и С-пептида, состоящего из 31 аминокислотного остатка. Инсулин и С-пептид в эквимолярных количествах включаются в секреторные гранулы. В гранулах инсулин соединяется с цинком, образуя димеры и гексамеры. Зрелые гранулы сливаются с плазматической мембраной, и инсулин и С-пептид секретируются во внеклеточную жидкость в результате экзоцитоза. После секреции в кровь олигомеры инсулина распадаются. Период полураспада инсулина в плазме крови составляет 3-10 минут, С-пептида - около 30 минут. Деградация инсулина происходит под действием фермента инсулиназы в основном в печени и в меньшей степени - в почках. Главным стимулятором синтеза и секреции инсулина является глюкоза. Секреция инсулина усиливается также некоторыми аминокислотами (особенно аргинином и лизином), кетоновыми телами и жирными кислотами. Адреналин, соматостатин и некоторые пептиды ЖКТ тормозят секрецию инсулина.
1 - синтез полипептидной цепи проинсулина; 2 - синтез происходит на полирибосомах, прикрепленных к наружной поверхности мембраны ЭР; 3 - сигнальный пептид отщепляется о завершении синтеза полипептидной цепи и образуется проинсулин; 4 - проинсулин транспортируется из ЭР в аппарат Гольджи и расщепляется на инсулин и С-пептид; 5 - инсулин и С-пептид включаются в секреторные гранулы и выделяются путем экзоцитоза (6); ЭР - эндоплазматический ретикулум; N - концевая часть молекулы; 3. Глюкагон - одноцепочечный полипептид, состоящий из 29 аминокислотных остатков. Биосинтез глюкагона происходит в α-клетках островков Лангерганса из неактивного предшественника препроглюкагона, который в результате частичного протеолиза превращается в активный гормон. Глюкоза и инсулин подавляют секрецию глюкагона; многие соединения, включая аминокислоты, жирные кислоты, нейромедиаторы (адреналин), ее стимулируют. Период полураспада гормона составляет ~5 минут. В печени глюкагон быстро разрушается под действием специфических протеаз. 4. Соматотропин синтезируется в виде прогормона в соматотрофных клетках, которые являются наиболее многочисленными в передней доле гипофиза. Гормон роста у всех видов млекопитающих представляет собой одноцепо- чечный пептид с молекулярной массой 22 кД, состоящий из 191 аминокислотного остатка и имеющий две внутримолекулярные дисульфидные связи. Секреция гормона роста носит пульсирующий характер с интервалами в 20-30 минут. Один из самых больших пиков отмечается вскоре после засыпания. Под влиянием различных стимулов (физические упражнения, голодание, белковая пища, аминокислота аргинин) даже у нерастущих взрослых людей уровень гормона роста в крови может возрастать до 30-100 нг/мл. Регуляция синтеза и секреции гормона роста осуществляется множеством факторов. Основной стимулирующий эффект оказывает соматолиберин, основной тормозящий - гипоталамический соматостатин. 5. Иодтиронины синтезируются в составе белка - тиреоглобулина (Тг) (рис. 11.5).
ЭР - эндоплазматический ретикулум; ДИТ - дийодтиронин; Тг - тиреоглобулин; Т3 - трийодтиронин, Т4 - тироксин. Тиреоглобулин синтезируется на рибосомах, далее поступает в комплекс Гольджи, а затем во внеклеточный коллоид, где он хранится и где происходит иодирование остатков тирозина. Образование йодтиронинов происходит в несколько этапов: транспорт иода в клетки щитовидной железы, окисление йода, йодирование остатков тирозина, образование йодтиронинов, транспорт йодтиронинов в кровь Тиреоглобулин - гликопротеин, содержит 115 остатков тирозина, синтезируется в базальной части клетки и хранится во внеклеточном коллоиде, где происходит йодирование остатков тирозина и образование йодтиронинов. Под действием тиреопероксидазы окисленный йод реагирует с остатками тирозина с образованием моноиодтиронинов (МИТ) и дийодтиронинов (ДИТ). Две молекулы ДИТ конденсируются с образованием Т4, а МИТ и ДИТ - с образованием Т3. Йодтиреоглобулин транспортируется в клетку путем эндоцитоза и гидролизуется ферментами лизосом с освобождением Т3 и Т4 (рис. 11.6).
Т3 является основной биологически активной формой йодтиронинов; его сродство к рецептору клеток-мишеней в 10 раз выше, чем у Т4. В периферических тканях в результате дейодирования части Т4 по пятому углеродному атому образуется так называемая «реверсивная» форма Т3, которая почти полностью лишена биологической активности В крови йодтиронины находятся в связанной форме в комплексе с тироксинсвязывающим белком. Только 0,03% Т4 и 0,3% Т3 находятся в свободном состоянии. Биологическая активность йодтиронинов обусловлена несвязанной фракцией. Транспортные белки служат своеобразным депо, которое может обеспечить дополнительное количество свободных гормонов. Синтез и секреция иодтиронинов регулируется гипоталамо-гипофизарной системой (рис. 11.7).
1 - тиреотропин-либерин стимулирует освобождение ТТГ; 2 - ТТГ стимулирует синтез и секрецию йодтиронинов; 3, 4 - иодтиронины тормозят синтез и секрецию ТТГ Йодтиронины регулируют процессы двух типов: • рост и дифференцировку тканей; • энергетический обмен. 6. Кортикостероиды. Общим предшественником всех кортикостероидов является холестерол. Источником холестерола для синтеза кортикостероидов служат его эфиры, поступающие в клетку в составе ЛПНП или депонированные в клетке. Освобождение холестерола из его эфиров и синтез кортикостероидов стимулируются кортикотропином. Реакции синтеза кортизола происходят в разных компартментах клеток коры надпочечников (см. рис. 11.12). При синтезе кортикостероидов образуется более 40 метаболитов, различающихся по структуре и биологической активности. Основными кортикостероидами, обладающими выраженной гормональной активностью, являются кортизол - главный представитель группы глюкокортикоидов, альдостерон - основной минералокортикоид и андрогены. На первом этапе синтеза кортикостероидов происходит превращение холестерола в прегненолон путем отщепления 6-углеродного фрагмента от боковой цепи холестерола и окисления углеродного атома С20. Прегненолон превращается в прогестерон - С21 предшественник стероидов - кортизола и альдостерона - и стероиды С19 - предшественники андрогенов. Каким именно стероидом окажется конечный продукт, зависит от набора ферментов в клетке и последовательности реакций гидроксилирования (рис. 11.8).
1 - превращение холестерола в прегненолон; 2 - образование прогестерона; 3 -гидроксилирование прогестерона (17-21-11) и образование кортизола; 4 - гидроксилирование прогестерона (21-11) и образование альдостерона; 5 - путь синтеза андрогенов Первичное гидроксилирование прогестерона 17-гидроксилазой, а затем 21- и 11-гидроксилазой приводит к синтезу кортизола. Реакции образования альдостерона включают гидроксилирование прогестерона сначала 21-гид- роксилазой, а затем 11-гидроксилазой (см. рис. 11.8). Скорость синтеза и секреции кортизола регулируется гипоталамо-гипофизарной системой по механизму обратной отрицательной связи (рис. 11.9). Стероидные гормоны транспортируются кровью в комплексе со специфическими транспортными белками. Катаболизм гормонов коры надпочечников происходит прежде всего в печени. Здесь протекают реакции гидроксилирования, окисления и
1 - стимуляция синтеза кортикотропин-либерина; 2 - кортикотропинлиберин стимулирует синтез и секрецию АКТГ; 3 - АКТГ стимулирует синтез и секрецию кортизола; 4 - кортизол тормозит секрецию АКТГ и кортиколиберина восстановления гормонов. Продукты катаболизма кортикостероидов (кроме кортикостерона и альдостерона) выводятся с мочой в форме 17-кетостероидов. Эти продукты метаболизма выделяются преимущественно в виде конъюгатов с глюкуроновой и серной кислотами. У мужчин 2/3 кетостероидов образуется за счет кортикостероидов и 1/3 - за счет тестостерона (всего 12-17 мг в сутки). У женщин 17-кетостероиды образуются преимущественно за счет кортикостероидов (7-12 мг в сутки). ТЕМА 11.4. РЕГУЛЯЦИЯ ОБМЕНА ОСНОВНЫХ ЭНЕРГОНОСИТЕЛЕЙ ПРИ НОРМАЛЬНОМ РИТМЕ ПИТАНИЯ 1. Энергетическая ценность основных пищевых веществ выражается в килокалориях и составляет: для углеводов - 4 ккал/г, для жиров - 9 ккал/г, для белков - 4 ккал/г. Взрослому здоровому человеку в сутки требуется 2000- 3000 ккал (8000-12 000 кДж) энергии. При обычном ритме питания промежутки между приемами пищи составляют 4-5 часов с 8-12-часовым ночным перерывом. В течение пищеварения и абсорбтивного периода (2-4 часа) основные энергоносители, используемые тканями (глюкоза, жирные кислоты, аминокислоты), могут поступать в кровь непосредственно из пищеварительного тракта. В постабсорбтивном периоде (промежуток времени после завершения пищеварения до следующего приема пищи) и при голодании энергетические субстраты образуются в процессе катаболизма депонированных энергоносителей. Основную роль в регуляции этих процессов играют инсулин и глюкагон. Антагонистами инсулина являются также адреналин, кортизол, иодтиронины и соматотропин (так называемые контринсулярные гормоны). Инсулин и контринсулярные гормоны обеспечивают баланс между потребностями и возможностями организма в получении энергии, необходимой для нормального функционирования и роста. Этот баланс определяется как энергетический гомеостаз. При нормальном ритме питания концентрация глюкозы в крови поддерживается на уровне 65-110 мг/дл (3,58-6,05 ммоль/л) благодаря влиянию двух основных гормонов - инсулина и глюкагона. Инсулин и глюкагон - главные регуляторы метаболизма при смене состояний пищеварения, постабсорбтивного периода и голодания. На периоды пищеварения приходится 10-15 час в сутки, а расход энергии происходит в течение 24 часов. Поэтому часть энергоносителей во время пищеварения запасается для использования в постабсорбтивном периоде. Печень, жировая ткань и мышцы - главные органы, обеспечивающие изменения метаболизма в соответствии с ритмом питания. Режим запасания включается после приема пищи и сменяется режимом мобилизации запасов после завершения абсорбтивного периода. 2. Изменения метаболизма основных энергоносителей в абсорбтивном периоде обусловлены, в основном, высоким инсулин-глюкагоновым индексом (рис. 11.10). Изменения метаболизма в печени. В печени увеличивается потребление глюкозы, что является следствием ускорения метаболических путей, в которых глюкоза превращается в депонируемые формы энергоносителей: гликоген и жиры. При повышении концентрации глюкозы в гепатоцитах происходит активация глюкокиназы, превращающей глюкозу в глюкозо-6-фосфат. Кроме этого, инсулин индуцирует синтез мРНК глюкокиназы. В результате повышается концентрация глюкозо-6-фосфата в гепатоцитах, что обусловливает ускорение синтеза гликогена. Этому также способствует одновременная инактивация гликогенфосфорилазы и активация гликогенсинтазы. Под влиянием инсулина в гепатоцитах ускоряется гликолиз в результате повышения активности и количества ключевых ферментов: глюкокиназы, фосфофруктокиназы и пируваткиназы. В то же время происходит торможение глюконеогенеза в результате инактивации фруктозо-1,6-бисфосфатазы и репрессии инсулином синтеза фосфоенолпируваткарбоксикиназы - ключевых ферментов глюконеогенеза (см. модуль 6). Повышение концентрации глюкозо-6-фосфата в гепатоцитах в абсорбтивном периоде сочетается с активным использованием NADPH для синтеза жирных кислот, что способствует стимуляции пентозофосфатного пути. Ускорение синтеза жирных кислот обеспечивается доступностью субстратов (ацетил-КоА и NADPH), образующихся при метаболизме глюкозы, а также активацией и индукцией ключевых ферментов синтеза жирных кислот инсулином.
1 - биосинтез гликогена в печени; 2 - гликолиз; 3 - биосинтез ТАГ в печени; 4 - биосинтез ТАГ в жировой ткани; 5 - биосинтез гликогена в мышцах; 6 - биосинтез белков в разных тканях, в том числе в печени; ЖК - жирные кислоты Аминокислоты, поступающие в печень из пищеварительного тракта, используются для синтеза белков и других азотсодержащих соединений, а их излишек либо поступает в кровь и транспортируется в другие ткани, либо дезаминируется с последующим включением безазотистых остатков в общий путь катаболизма (см. модуль 9). Изменения метаболизма в адипоцитах. Основная функция жировой ткани - запасание энергоносителей в форме триацилглицеролов. Под влиянием инсулина ускоряется транспорт глюкозы в адипоциты. Повышение внутриклеточной концентрации глюкозы и активация ключевых ферментов гликолиза обеспечивают образование ацетил-КоА и глицерол-3-фосфата, необходимых для синтеза ТАГ. Стимуляция пентозофосфатного пути обеспечивает образование NADPH, необходимого для синтеза жирных кислот. Однако биосинтез жирных кислот de novo в жировой ткани человека протекает с высокой скоростью только после предшествующего голодания. При нормальном ритме питания для синтеза ТАГ используются в основном жирные кислоты, поступающие из хиломикронов и ЛПОНП под действием ЛП-липазы (см. модуль 8). Так как гормончувствительная ТАГ-липаза в абсорбтивном состоянии находится в дефосфорилированной, неактивной форме, процесс липолиза тормозится. Изменение метаболизма в мышцах. Под влиянием инсулина ускоряется транспорт глюкозы в клетки мышечной ткани. Глюкоза фосфорилируется и окисляется для обеспечения клеток энергией, а также используется для синтеза гликогена. Жирные кислоты, поступающие из хиломикронов и ЛПОНП, в этот период играют незначительную роль в энергетическом обмене мышц. Поток аминокислот в мышцы и биосинтез белков также возрастает под влиянием инсулина, особенно после приема белковой пищи и в период мышечной работы. 3. Изменения метаболизма основных энергоносителей при смене абсорбтивного состояния на постабсорбтивное. В постабсорбтивном периоде при снижении инсулин-глюкагонового индекса изменения метаболизма направлены главным образом на поддержание концентрации в крови глюкозы, которая служит главным энергетическим субстратом для мозга и единственным источником энергии для эритроцитов. Основные изменения метаболизма в этот период происходят в печени и жировой ткани (рис. 11.11) и направлены на пополнение глюкозы за счет внутренних резервов и на использование других энергетических субстратов (жиров и аминокислот). Изменения метаболизма в печени. Под влиянием глюкагона ускоряется мобилизация гликогена (см. модуль 6). Запасы гликогена в печени истощаются в течение 18-24-часового голодания. Главным источником глюкозы по мере исчерпания запасов гликогена становится глюконеогенез, который начинает ускоряться через 4-6 часов после последнего приема пищи. Субстратами для синтеза глюкозы служат лактат, глицерол и аминокислоты. Скорость синтеза жирных кислот снижается вследствие фосфорилирования и инактивации ацетил-КоА-карбоксилазы при фосфорилировании, а скорость β-окисления возрастает. Вместе с тем увеличивается снабжение печени жирными кислотами, которые транспортируются из жировых депо в результате ускорения липолиза. Ацетил-КоА, образующийся при окислении жирных кислот, используется в печени для синтеза кетоновых тел. В жировой ткани снижается скорость синтеза ТАГ и стимулируется липолиз. Стимуляция липолиза является результатом активации гормончувствительной ТАГ-липазы адипоцитов под влиянием глюкагона. Жирные кислоты становятся важными источниками энергиии в печени, мышцах и жировой ткани. Таким образом, в постабсорбтивном периоде концентрация глюкозы в крови поддерживается на уровне 60-100 мг/дл (3,5-5,5 ммоль/л), а уровень жирных кислот и кетоновых тел возрастает.
I - снижение инсулин-глюкагонового индекса; 2 - распад гликогена; 3, 4 - транспорт глюкозы в мозг и эритроциты; 5 - катаболизм жиров; 6 - транспорт жиров в печень и мышцы; 7 - синтез кетоновых тел в печени; 8 - транспорт кетоновых тел в мышцы; 9 - глюконеогенез из аминокислот; 10 - синтез и выведение мочевины; II - транспорт лактата в печень и включение в глюконеогенез; 12 - глюконеогенез из глицерола; КТ -кетоновые тела; ЖК - жирные кислоты ТЕМА 11.5. ИЗМЕНЕНИЯ МЕТАБОЛИЗМА ПРИ ГИПО- И ГИПЕРСЕКРЕЦИИ ГОРМОНОВ Изменение скорости синтеза и секреции гормонов может происходить не только как адаптационный процесс, возникающий в ответ на изменение физиологической активности организма, но часто является результатом нарушений функциональной активности эндокринных желез при развитии в них патологических процессов или нарушений регуляции. Эти нарушения могут проявляться либо в форме гипофункции, приводящей к снижению количества гормона, либо гиперфункции, сопровождающейся избыточным его синтезом. 1. Гиперфункция щитовидной железы (гипертиреоз) проявляется в нескольких клинических формах. Диффузный токсический зоб (базедова болезнь, болезнь Грейвса) - наиболее распространенное заболевание щитовидной железы. При этом заболевании отмечается увеличение размеров щитовидной железы (зоб), повышение концентрации йодтиронинов в 2-5 раз и развитие тиреотоксикоза. Характерными признаками тиреотоксикоза являются увеличение основного обмена, учащение сердцебиений, мышечная слабость, потеря массы тела (несмотря на повышенный аппетит), потливость, повышение температуры тела, тремор и экзофтальм (пучеглазие). Эти симптомы отражают одновременную стимуляцию йодтиронинами как анаболических (рост и дифференцировка тканей), так и катаболических процессов (катаболизм углеводов, липидов и беков). В большей мере усиливаются процессы катаболизма, о чем свидетельствует отрицательный азотистый баланс. Гипертиреоз может возникать в результате различных причин: развития опухоли, воспаления (тиреоидит), избыточного поступления йода и йодсодержащих препаратов, аутоиммунных реакций. Аутоиммунный гипертиреоз возникает в результате образования антител к рецепторам тиреотропного гормона в щитовидной железе. Один из них - иммуноглобулин (IgG) - имитирует действие тиреотропина, взаимодействуя с рецепторами ТТГ на мембране клеток щитовидной железы. Это приводит к диффузному разрастанию щитовидной железы и избыточной неконтролируемой продукции Т3 и Т4, поскольку образование IgG не регулируется по механизму обратной связи. Уровень ТТГ при этом заболевании снижен вследствие подавления функции гипофиза высокими концентрациями йодтиронинов. 2. Гипотиреоз может быть результатом недостаточного поступления йода в организм - эндемического зоба. Реже гипотиреоз возникает в результате врожденных дефектов ферментов, участвующих в синтезе (например, тиреопероксиразы) йодтиронинов, или как осложнение других болезней, при которых повреждаются гипоталамус, гипофиз или щитовидная железа. При некоторых формах гипотиреоза в крови обнаруживаются антитела к тиреоглобулину. Гипофункция щитовидной железы в раннем детском возрасте приводит к задержке физического и умственного развития - кретинизм. У взрослых гипофункция проявляется как микседему (слизистый отек). Главным проявлением микседемы является избыточное накопление в коже протеогликанов и воды. Основные симптомы гипотиреоза: сонливость, снижение толерантности к холоду, увеличение массы тела, снижение температуры тела. 3. Гиперкортицизм. Избыточное образование кортикостероидов, главным образом кортизола, - гиперкортицизм - часто является результатом нарушения регуляторных механизмов синтеза кортизола: • при опухоли гипофиза и повышенной продукции кортикотропина (болезнь Иценко-Кушинга); • при опухолях надпочечников, продуцирующих кортизол (синдром Иценко-Кушинга). Главные проявления гиперкортицизма: гиперглюкоземия и снижение толерантности к глюкозе вследствие стимуляции глюконеогенеза и гипертензия как результат проявления минералокортикоидной активности кортизола и повышения концентрации ионов Na+. 4. Гипокортицизм. Наследственная адреногенитальная дистрофия в 95% случаев является следствием дефицита 21-гидроксилазы (см. рис. 11.8). При этом увеличивается образование 17-ОН прогестерона и продукции андрогенов. Характерными симптомами заболевания являются раннее половое созревание у мальчиков и развитие мужских половых признаков у девочек. При частичной недостаточности 21-гидроксилазы у женщин может нарушаться менструальный цикл. Приобретенная недостаточность надпочечников может развиваться в результате туберкулезного или аутоиммунного повреждения клеток коры надпочечников и снижения синтеза кортикостероидов. Потеря регуляторного контроля со стороны надпочечников приводит к повышению секреции кортикотропина. В этих случаях у больных отмечается усиление пигментации кожи и слизистых (аддисонова болезнь), что обусловлено повышенной продукцией кортикотропина и других производных ПОМК, в частности меланоцитстимулирующего гормона (см. рис. 11.3). Основные клинические проявления надпочечниковой недостаточности: гипотензия, мышечная слабость, гипонатриемия, потеря массы тела, непереносимость стресса. Недостаточность функции коры надпочечников часто является следствием длительного применения кортикостероидных препаратов, подавляющих синтез кортикотропина по механизму обратной связи. Отсутствие стимулирующих сигналов приводит к атрофии клеток коры надпочечников. При резкой отмене гормональных препаратов может развиться острая надпочечниковая недостаточность (так называемый синдром «отмены»), которая представляет большую угрозу для жизни, так как сопровождается декомпенсацией всех видов обмена и процессов адаптации. Она проявляется сосудистым коллапсом, резкой адинамией, потерей сознания. Такое состояние возникает вследствие нарушения обмена электролитов, которое приводит к потере ионов Na+ и С1- с мочой и обезвоживанию за счет потери внеклеточной жидкости. Изменение углеводного обмена проявляется в снижении уровня сахара в крови, уменьшении запаса гликогена в печени и скелетных мышцах. ЗАДАНИЯ ДЛЯ ВНЕАУДИТОРНОЙ РАБОТЫ 1. Перенесите в тетрадь и заполните табл. 11.1. Таблица 11.1. Инсулин и основные контринсулярные гормоны
2. Используя рис. 11.4, выпишите этапы синтеза инсулина. Объясните, какие причины могут привести к развитию инсулиновой недостаточности? Почему в этих случаях с целью диагностики можно определять в крови концентрацию С-пептида? 3. Изучите схему синтеза йодтиронинов (рис. 11.5). Опишите основные этапы их синтеза и нарисуйте схему регуляции синтеза и секреции тиреоидных гормонов. Объясните основные проявления гипо- и гипертиреоза. Почему при применении тироксина как лекарства необходим постоянный контроль уровня ТТГ в крови? 4. Изучите последовательность этапов синтеза кортизола (рис. 11.8). Найдите на схеме этапы, катализируемые ферментами, дефект которых является причиной возникновения адреногенитального синдрома. 5. Опишите схему внутриклеточного цикла синтеза кортизола начиная с взаимодействия АКТГ с рецептором (рис. 11.12), заменив цифры названиями участвующих в них белков. 6. Нарисуйте схему регуляции синтеза и секреции кортикостероидов. Объясните причины и проявления синдрома отмены стероидных препаратов. 7. Опишите последовательность событий, которые приводят к повышению концентрации глюкозы в крови в течение первого часа после приема пищи и ее последующему возвращению к исходному уровню в течение 2 часов (рис. 11.13). Объясните роль гормонов в этих событиях. 8. Проанализируйте изменения гормонального статуса и метаболизма в печени, жировой ткани и мышцах в абсорбтивном (рис. 11.10) и постабсорбтивном периодах (рис. 11.11). Назовите процессы, обозначенные цифрами. Укажите регуляторные ферменты и механизм изменения их активности, учитывая, что первичным сигналом для стимуляции этих процессов является изменение концентрации глюкозы в крови и реципрокные изменения концентрации инсулина и глюкагона (рис. 11.11).
ЭХС - эфиры холестерола; ХС - холестерол ЗАДАНИЯ ДЛЯ САМОКОНТРОЛЯ 1. Выберите правильные ответы. Гормоны: A. Проявляют свои эффекты через взаимодействие с рецепторами Б. Синтезируются в задней доле гипофиза B. Измененяют активность ферментов путем частичного протеолиза Г. Индуцируют синтез ферментов в клетках-мишенях Д. Синтез и секреция регулируется по механизму обратной связи
2. Выберите правильный ответ. Глюкагон в жировой ткани активирует: A. Гормончувствительную ТАГ-липазу Б. Глюкозо-6-фосфатдегидрогеназу B. Ацетил-КоА-карбоксилазу Г. ЛП-липазу Д. Пируваткиназу 3. Выберите правильные ответы. Йодтиронины: A. Синтезируются в гипофизе Б. Взаимодействуют с внутриклеточными рецепторами B. Стимулируют работу Na, Ka-АТФазы Г. В высоких концентрациях ускоряют процессы катаболизма Д. Участвуют в ответной реакции на охлаждение 4. Установите соответствие: A. Базедова болезнь Б. Микседема B. Эндемический зоб Г. Кретинизм Д. Аутоиммунный тиреоидит 1. Возникает при гипофункции щитовидной железы в раннем возрасте 2. Сопровождается накоплением протеогликанов и воды в коже 3. Является следствием образования иммуноглобулина, имитирующего действие ТТГ 5. Выберите правильные ответы. Для абсорбтивного периода характерно: A. Повышение концентрации инсулина в крови Б. Ускорение синтеза жиров в печени B. Ускорение глюконеогенеза Г. Ускорение гликолиза в печени Д. Повышение концентрации глюкагона в крови 6. Выберите правильные ответы. Под влиянием инсулина в печени ускоряются: A. Биосинтез белков Б. Биосинтез гликогена B. Глюконеогенез Г. Биосинтез жирных кислот Д. Гликолиз 7. Установите соответствие. Гормон: A. Инсулин Б. Глюкагон B. Кортизол Г. Адреналин Д. ТТГ Функция: 1. Стимулирует синтез жиров из глюкозы в печени 2. Стимулирует мобилизацию гликогена в мышцах 3. Стимулирует синтез йодтиронинов 8. Выберите правильные ответы. Стероидные гормоны: A. Проникают в клетки-мишени Б. Транспортируются кровью в комплексе со специфическими белками B. Стимулируют реакции фосфорилирования белков Г. Взаимодействуют с хроматином и изменяют скорость транскрипции Д. Участвуют в процессе трансляции. 9. Выберите правильные ответы. Инсулин: A. Ускоряет транспорт глюкозы в мышцы Б. Ускоряет синтез гликогена в печени B. Стимулирует липолиз в жировой ткани Г. Ускоряет глюконеогенез Д. Ускоряет транспорт глюкозы в адипоциты ЭТАЛОНЫ ОТВЕТОВ К «ЗАДАНИЯМ ДЛЯ САМОКОНТРОЛЯ» 1. А, Г, Д 6. А, Б, Г, Д 2. А 7. 1-А, 2-Г, 3-Д 3. Б, В, Г, Д 8. А, Б, Г 4. 1-Г, 2-Б, 3 - А 9. А, Б, Д 5. А, Б, Г ОСНОВНЫЕ ТЕРМИНЫ И ПОНЯТИЯ 1. Гормон 2. Препрогормон 3. Стимулы для синтеза и секреции 4. Клетки-мишени 5. Рецепоры 6. Иерархия регуляторных систем 7. Аутокринный механизм действия 8. Паракринный механизм действия 9. Гомеостаз 10. Абсорбтивный период 11. Постабсорбтивный период 12. Адаптация 13. Гипофункция 14. Гиперфункия 15. Контринсулярные гормоны ЗАДАНИЯ ДЛЯ АУДИТОРНОЙ РАБОТЫ Решите задачи 1. При обследовании больных с явлениями гиперкортицизма используют функциональную пробу с «нагрузкой» дексаметазоном (дексаметазон - структурный аналог кортизола). Как изменится концентрация 17-кетостероидов в моче пациентов после введения дексаметазона, если причиной гиперкортицизма является: а) гиперпродукция кортикотропина; б) гормонально активная опухоль надпочечников. 2. Родители пятилетней девочки обратились в медицинский центр для консультации. При обследовании у ребенка отмечены проявления вторичных мужских половых признаков: гипертрофия мышц, избыточное оволосенение, снижение тембра голоса. В крови повышен уровень АКТГ. Врач диагностировал адреногенитальный синдром (врожденная дисфункция коры надпочечников). Обоснуйте диагноз врача. Для этого: а) представьте схему синтеза стероидных гормонов; назовите основные физиологически активные кортикостероиды и укажите их функции; б) назовите ферменты, недостаточность которых является причиной описанных выше симптомов; в) укажите образование каких продуктов синтеза кортикостероидов увеличивается при этой патологии; г) объясните, почему у ребенка в крови повышена концентрация АКТГ. 3. Одна из форм болезни Аддисона является следствием атрофии клеток коры надпочечников при длительном лечении кортикостероидными препаратами. Основные проявления болезни: мышечная слабость, гипоглюкоземия, дистрофические изменения в мышцах, снижение артериального давления; в ряде случаев у таких больных отмечается усиление пигментации кожи и слизистых. Как объяснить перечисленные симптомы заболевания? Для объяснения: а) представьте схему синтеза стероидных гормонов; назовите основные физиологически активные кортикостероиды и укажите их функции; б) укажите, дефицит каких кортикостероидов является причиной гипоглюкоземии и мышечной дистрофии при этом заболевании; в) назовите причину повышенной пигментации кожи при болезни Аддисона. 4. Пациенту N с гипотиреозом врач назначил лечение, включающее прием тироксина. Спустя 3 месяца после начала лечения уровень ТТГ в крови снизился незначительно. Почему этому больному врач рекомендовал увеличить дозу тироксина? Для ответа: а) представьте в виде схемы механизм регуляции синтеза и секреции тиреоидных гормонов; б) используя схему, обоснуйте рекомендацию врача. 5. Девушка 18-лет живущая в горном селении, обратилась к эндокринологу с жалобами на общую слабость, снижение температуры тела, ухудшение настроения. Пациентка была направлена на анализ крови на ТТГ и йодтиронины. Результаты анализа показали повышение коцентрации ТТГ и снижение концентрации Т4.. Объясните: а) какое заболевание можно предположить у пациентки; б) что может быть причиной такой патологии; в) есть ли связь между местом проживания и возникновением данного заболевания; г) какую диету следует соблюдать в целях профилактики этой патологии; д) схему регуляции синтеза йодтиронинов и результаты анализа крови у обследуемой. 6. Для лечения диффузного токсического зоба применяют тиреостатические препараты группы тионамидов (тиамазол). Механизм действия тионамидов заключается в том, что, попадая в щитовидную железу, они подавляют активность тиреопероксидазы. Объясните результат лечебного действия тионамидов. Для этого: а) назовите основные причины и клинические проявления тиреотоксикоза; б) приведите схему синтеза йодтиронинов и укажите этапы, на которые действуют лекарства; в) укажите, как изменится концентрация йодтиронинов и ТТГ в результате лечения; г) опишите изменения метаболизма при лечении тионамидами. Модульная единица 2 БИОХИМИЧЕСКИЕ ИЗМЕНЕНИЯ ОБМЕНА ВЕЩЕСТВ ПРИ ГОЛОДАНИИ И САХАРНОМ ДИАБЕТЕ Цели изучения Уметь: 1. Интерпретировать изменения в метаболизме углеводов, жиров и белков при голодании и физической нагрузке как результат действия контринсулярных гормонов. 2. Анализировать молекулярные механизмы причин сахарного диабета. 3. Объяснять механизмы возникновения симптомов сахарного диабета как следствие изменений скоростей метаболических процессов. 4. Интерпретировать основные различия в обмене веществ при голодании и сахарном диабете. Знать: 1. Изменения гормонального статуса при голодании. 2. Изменение обмена основных энергоносителей при голодании. 3. Изменение гормонального статуса и энергетического метаболизма при сахарном диабете. 4. Основные симптомы сахарного диабета и механизмы их возникновения. 5. Патогенез острых осложнений при диабете. 6. Биохимические основы поздних осложнений сахарного диабета. 7. Подходы к лабораторной диагностике сахарного диабета. 8. Молекулярные механизмы принципов лечения сахарного диабета и перспективные направления лечения. ТЕМА 11.6. ИЗМЕНЕНИЯ ГОРМОНАЛЬНОГО СТАТУСА И МЕТАБОЛИЗМА ПРИ ГОЛОДАНИИ И ФИЗИЧЕСКОЙ РАБОТЕ 1. В постабсорбтивном периоде и голодании уровень глюкозы в плазме крови падает до нижней границы нормы. Отношение инсулин - глюкагон снижается. В этих условиях возникает состояние, для которого характерно преобладание процессов катаболизма жиров, гликогена и белков на фоне общего снижения скорости метаболизма. Под влиянием контринсулярных гормонов в этот период происходит обмен субстратами между печенью, жировой тканью, мышцами и мозгом. Этот обмен служит двум целям: • поддержанию концентрации глюкозы в крови за счет глюконеогенеза для обеспечения глюкозозависимых тканей (мозга, эритроцитов); • мобилизации других «топливных» молекул, в первую очередь жиров, для обеспечения энергией всех других тканей. Проявление этих изменений позволяет условно выделить три фазы голодания. Вследствие переключения метаболизма на режим мобилизации энергоносителей даже после 5-6 недель голодания концентрация глюкозы в крови составляет не менее 65 мг/дл. Основные изменения при голодании происходят в печени, жировой ткани и мышцах (рис. 11.14). 2. Фазы голодания. Голодание может быть кратковременным - в течение суток (первая фаза), продолжаться в течение недели (вторая фаза) или нескольких недель (третья фаза). В первую фазу концентрация инсулина в крови снижается примерно в 10-15 раз по сравнению с периодом пищеварения, а концентрация глюкагона и кортизола увеличивается. Запасы гликогена исчерпываются, нарастает скорость мобилизации жиров и скорость глюконеогенеза из аминокислот и глицерола, концентрация глюкозы в крови снижается до нижней границы нормы (60 мг/дл).
1 - снижение инсулин-глюкогонового индекса; 2 - мобилизация гликогена; 3, 4 - транспорт ГЛК в мозг и эритроциты; 5 - мобилизация ТАГ; 6 - транспорт ЖК в мышцы; 7 - синтез кетоновых тел; 8 - транспорт ЖК в печени; 9 - транспорт АК в печень; 10 - глюконеогенез из АК; 11 - транспорт лактата в печень; 12 - транспорт глицерола в печень. Пунктиром обозначены процессы, скорость которых снижается Во вторую фазу продолжается мобилизация жиров, концентрация жирных кислот в крови повышается, увеличивается скорость образования кетоновых тел в печени и, соответственно, их концентрация в крови; ощущается запах ацетона, который выделяется с выдыхаемым воздухом и потом от голодающего человека. Глюконеогенез продолжается за счет распада тканевых белков. В третью фазу снижается скорость распада белков и скорость глюконеогенеза из аминокислот. Скорость метаболизма замедляется. Азотистый баланс во все фазы голодания отрицательный. Для мозга важным источником энергии, наряду с глюкозой, становятся кетоновые тела. 3. Изменения метаболизма основных энергоносителей при голодании. Обмен углеводов. Запасы гликогена в организме истощаются в течение 24-часового голодания. Таким образом, за счет мобилизации гликогена обеспечивается только кратковременное голодание. Основной процесс, обеспечивающий ткани глюкозой в период голодания, - глюконеогенез. Глюконеогенез начинает ускоряться через 4-6 часов после последнего приема пищи и становится единственным источником глюкозы в период длительного голодания. Основные субстраты глюконеогенеза - аминокислоты, глицерол и лактат. 4. Обмен жиров и кетоновых тел. Основным источником энергии в первые дни голодания становятся жирные кислоты, которые образуются из ТАГ в жировой ткани. В печени ускоряется синтез кетоновых тел. Синтез кетоновых тел начинается в первые дни голодания. Используются кетоновые тела в основном в мышцах. Энергетические потребности мозга частично обеспечиваются также кетоновыми телами. После 3 недель голодания в мышцах снижается скорость окисления кетоновых тел и мышцы почти исключительно используют жирные кислоты. Концентрация кетоновых тел в крови возрастает. Использование кетоновых тел мозгом продолжается, но становится менее активным из-за снижения скорости глюконеогенеза и снижения концентрации глюкозы. 5. Обмен белков. В течение нескольких первых дней голодания быстро распадаются мышечные белки - основной источник субстратов для глюконеогенеза. После нескольких недель голодания скорость глюконеогенеза из аминокислот снижается в основном вследствие снижения потребления глюкозы и использования кетоновых тел в мозге. Снижение скорости глюконеогенеза из аминокислот необходимо для сбережения белков, так как потеря 1/3 всех белков может привести к смерти. Продолжительность голодания зависит от того, как долго могут синтезироваться и использоваться кетоновые тела. Однако для окисления кетоновых тел необходим оксалоацетат и другие компоненты ЦТК. В норме они образуется из глюкозы и аминокислот, а при голодании только из аминокислот. ТЕМА 11.7. ИЗМЕНЕНИЯ ГОРМОНАЛЬНОГО СТАТУСА И МЕТАБОЛИЗМА ПРИ САХАРНОМ ДИАБЕТЕ 1. Сахарный диабет возникает вследствие относительного или абсолютного дефицита инсулина. В соответствии с классификацией ВОЗ различают две основные формы заболевания: диабет I типа - инсулинзависимый (ИЗСД), и диабет II типа (ИНСД) - инсулиннезависимый. 2. ИЗСД является следствием разрушения β-клеток островков Лангерганса в результате аутоиммунных реакций. Провоцировать возникновение диабета I типа может вирусная инфекция, вызывающая деструкцию β-клеток. К таким вирусам относятся вирусы оспы, краснухи, кори, цитомегаловирус, эпидимического паротита, вирус Коксаки, аденовирус. На долю ИЗСД приходится примерно 25-30% всех случаев диабета. Как правило, разрушение β-клеток происходит медленно и начало заболевания не сопровождается нарушениями метаболизма. Когда погибает 80-95% клеток, возникает абсолютный дефицит инсулина и развиваются тяжелые метаболические нарушения. ИЗСД поражает в большинстве случаев детей, подростков и молодых людей, но может проявиться в любом (начиная с годовалого) возрасте. 3. ИНСД развивается вследствие нарушения превращения проинсулина в инсулин, регуляции секреции инсулина, повышения скорости катаболизма инсулина, повреждения механизмов передачи инсулинового сигнала в клетки-мишени (например, дефекта рецептора инсулина, повреждения внутриклеточных посредников инсулинового сигнала и т.д.), образование антител к рецепторам инсулина, причем концентрация инсулина в крови может быть нормальной или даже повышенной. К факторам, определяющим развитие и клиническое течение болезни, относятся ожирение, неправильный режим питания, малоподвижный образ жизни, стресс. ИНСД поражает людей, как правило, старше 40 лет, развивается постепенно, симптомы выражены умеренно. Острые осложнения бывают редко. 4. Изменения метаболизма при сахарном диабете. При сахарном диабете, как правило, соотношение инсулин - глюкагон снижено. При этом ослабевает стимуляция процессов депонирования гликогена и жиров и усиливается мобилизация запасов энергоносителей. Печень, мышцы и жировая ткань даже после приема пищи функционируют в режиме постабсорбтивного состояния. 5. Симптомы сахарного диабета. Гиперглюкоземия. Для всех форм диабета характерно повышение концентрации глюкозы в крови - гиперглюкоземия, как после приема пищи, так и натощак, а также глюкозурия. После приема пищи концентрация глюкозы может достигать 300-500 мг/дл и сохраняется на высоком уровне в постабсорбтивном периоде, т.е. снижается толерантность к глюкозе. Снижение толерантности к глюкозе наблюдается и в случаях скрытой (латентной) формы сахарного диабета. В этих случаях у людей отсутствуют жалобы и клинические симптомы, характерные для сахарного диабета, а концентрация глюкозы в крови натощак соответствует верхней границе нормы. Однако использование провокационных проб (например, сахарной нагрузки) выявляет снижение толерантности к глюкозе (рис. 11.15). Повышение концентрации глюкозы при ИЗСД в плазме крови обусловлено несколькими причинами. При снижении инсулин-глюкагонового индекса усиливаются эффекты контринсулярных гормонов, уменьшается количество белков - переносчиков глюкозы (ГЛЮТ-4) на мембранах инсулинзависимых клеток (жировой ткани и мышц). Следовательно, снижается потребление глюкозы этими клетками. В мышцах и печени глюкоза не депонируется в виде гликогена, в жировой ткани уменьшается скорость синтеза и депонирования жиров. Кроме того, действие конринсулярных гормонов, в первую очередь глюкагона, активирует глюконеогенез из аминокислот, глицерола и лактата. Повышение уровня глюкозы в крови при сахарном диабете выше концентрационного почечного порога, равного 180 мг/дл, становится причиной выделения глюкозы с мочой. Кетонемия является характерным признаком сахарного диабета. При низком соотношении инсулин - глюкагон жиры не депонируются, ускоряется их катаболизм, так как гормончувствительная липаза в жировой ткани находится в фосфорилированной активной форме. Концентрация неэтерифицированных жирных кислот в крови повышается. Печень захватывает жирные кислоты и окисляет их до ацетил-КоА, который в свою очередь
Определение толерантности к глюкозе используют для диагностики сахарного диабета. Обследуемый принимает раствор глюкозы из расчета 1 г на 1 кг массы тела (сахарная нагрузка). Концентрацию глюкозы в крови измеряют в течение 2-3 часов с интервалами в 30 минут. 1 - у здорового человека, 2 - у больного сахарным диабетом превращается в β-гидроксимасляную и ацетоуксусную кислоты, в результате чего в крови повышается концентрация кетоновых тел - кетонемия. В тканях ацетоацетат частично декарбоксируется до ацетона, запах которого исходит от больных сахарным диабетом и ощущается даже на расстоянии. Увеличение концентрации кетоновых тел в крови (выше 20 мг/дл, иногда до 100 мг/дл) приводит к кетонурии. Накопление кетоновых тел снижает буферную емкость крови и вызывает ацидоз (кетоацидоз). Гиперлипопротеинемия. Пищевые жиры не депонируются в жировой ткани вследствие ослабления процессов запасания и низкой активности ЛП-липазы, а поступают в печень, где превращаются в триацилглицеролы, которые транспортируются из печени в составе ЛПОНП. Азотемия. При диабете дефицит инсулина приводит к снижению скорости синтеза и усилению распада белков в организме. Это вызывает повышение концентрации аминокислот в крови. Аминокислоты поступают в печень и дезаминируются. Безазотистые остатки гликогенных аминокислот включаются в глюконеогенез, что еще более усиливает гиперглюкоземию. Образующийся при этом аммиак вступает в орнитиновый цикл, что приводит к увеличению концентрации мочевины в крови и соответственно в моче - азотемии и азотурии. Полиурия. Для выведения большого количества глюкозы, кетоновых тел и мочевины требуется большой объем жидкости, в результате чего может наступить обезвоживание организма. Это объясняется особенностями концентрационной способности почек. Например, выделение мочи у больных возрастает в несколько раз и в некоторых случаях достигает 8-9 л в сутки, но чаще не превышает 3-4 л. Этот симптом называется полиурией. Потеря воды вызывает постоянную жажду и увеличение потребления воды - полидипсию. 6. Острые осложнения сахарного диабета. Механизмы развития диабетической комы. Нарушения обмена углеводов, жиров и белков при сахарном диабете могут приводить к развитию коматозных состояний (острые осложнения). Диабетическая кома проявляется как резкое нарушение всех функций организма, сопровождающееся потерей сознания. Основными предшественниками диабетической комы являются ацидоз и дегидратация тканей (рис. 11.16). При декомпенсации диабета развивается нарушение водно-электролитного обмена. Причиной этого является гиперглюкоземия, сопровождающаяся повышением осмотического давления в сосудистом русле. Для сохранения осмолярности начинается компенсаторное перемещение жидкости из клеток и внеклеточного пространства в сосудистое русло. Это ведет к потере тканями воды и электролитов, прежде всего ионов Na+, K+, Cl-, HCO3-. В результате развиваются тяжелая клеточная дегидратация и дефицит внутриклеточных ионов (прежде всего К+), сопровождающаяся общей дегидратацией. Это приводит к снижению периферического кровообращения, уменьшению мозгового и почечного кровотока и гипоксии. Диабетическая кома развивается медленно, в течение нескольких дней, но иногда может
возникнуть и в течение нескольких часов. Первыми признаками могут быть тошнота, рвота, заторможенность. Артериальное давление у больных снижено. Коматозные состояния при сахарном диабете могут проявляться в трех основных формах: кетоацидотической, гиперосмолярной и лактоацидотической. Для кетоацидотической комы характерны выраженный дефицит инсулина, кетоацидоз, полиурия, полидипсия. Гиперглюкоземия (20-30 ммоль/л), обусловленная инсулиновой недостаточностью, сопровождается большими потерями жидкости и электролитов, дегидратацией и гиперосмолярностью плазмы. Общая концентрация кетоновых тел достигает 100 мг/дл и выше. При гиперосмолярной коме наблюдается чрезвычайно высокие уровни глюкозы в плазме крови, полиурия, полидипсия, всегда проявляется тяжелая дегидратация. Предполагают, что у большинства больных гиперглюкоземия обусловлена сопутствующим нарушением функции почек. Кетоновые тела в сыворотке крови обычно не определяются. При лактоацидотической коме преобладающими являются гипотония, снижение периферического кровообращения, гипоксия тканей, приводящая к смещению метаболизма в сторону анаэробного гликолиза, что обусловливает повышение концентрации молочной кислоты в крови (лактоацидоз). 7. Поздние осложнения сахарного диабета являются следствием длительной гиперглюкоземии и часто приводят к ранней инвалидизации больных. Гиперглюкоземия приводит к повреждению кровеносных сосудов и нарушению функций различных тканей и органов. Одним из основных механизмов повреждения тканей при сахарном диабете является глюкозилирование белков и связанное с этим нарушение функции клеток тканей, изменение реологических свойств крови и гемодинамики (текучесть, вязкость). Некоторые соединения в норме содержат углеводные компоненты (гликопротеины, протеогликаны, гликолипиды). Синтез этих соединений происходит в результате ферментативных реакций (ферментативное глюкозилирование). Однако в организме человека может происходить и неферментативное взаимодействие альдегидной группы глюкозы со свободными аминогруппами белков (неферментативное глюкозилирование). В тканях здоровых людей этот процесс протекает медленно, а при гиперглюкоземии ускоряется. Одним из первых признаков диабета является увеличение в 2-3 раза глюкозилированного гемоглобина. На протяжении всего срока существования эритроцитов глюкоза свободно проникает через его мембрану и без участия ферментов необратимо связывается с гемоглобином, преимущественно β-цепями. При этом образуется глюкозилированная форма гемоглобина HbA1c. Эта форма гемоглобина в небольшом количестве имеется и у здоровых людей. В условиях хронической гиперглюкоземии процент HbA1c по отношению к общему количеству гемоглобина увеличивается. Степень глюкозилирования белков зависит от скорости их обновления. В медленно обменивающихся белках накапливается больше изменений. К медленно обменивающимся белкам относятся белки межклеточного матрикса, базальных мембран, хрусталика глаза (кристаллины). Утолщение базальных мембран - один из ранних и постоянных признаков сахарного диабета, проявляющихся в форме диабетических ангиопатий. Изменения, проявляющиеся в снижении эластичности артерий, поражении крупных и средних сосудов мозга, сердца, нижних конечностей, называются диабетическими макроангиопатиями. Они развиваются вследствие глюкозилирования белков межклеточного матрикса - коллагена и эластина, что приводит к снижению эластичности сосудов и нарушению кровообращения. Результат повреждения капилляров и мелких сосудов - микроангиопатии проявляются в форме нефро- и ретинопатии. Причиной некоторых поздних осложнений сахарного диабета (катаракты, ретинопатии) может быть повышение скорости превращения глюкозы в сорбитол. Сорбитол не используется в других метаболических путях, а скорость его диффузии из клеток невелика. У больных сахарным диабетом сорбитол накапливается в сетчатке и хрусталике глаза, клетках клубочков почек, шванновских клетках, в эндотелии. Сорбитол в высоких концентрациях токсичен для клеток. Его накопление в нейронах приводит к увеличению осмотического давления, набуханию клеток и отеку тканей. Помутнение хрусталика, или катаракта, может развиться как вследствие вызванного накоплением сорбитола набухания хрусталика и нарушения упорядоченной структуры кристаллинов, так и вследствие глюкозилирования кристаллинов, которые образуют многомолекулярные агрегаты, увеличивающие преломляющую способность хрусталика. ЗАДАНИЯ ДЛЯ ВНЕАУДИТОРНОЙ РАБОТЫ 1. Рассмотрите рис. 11.14, изобразите схемы процессов, которые ускоряются в печени и других тканях при наступлении постабсорбтивного периода, выпишите названия метаболических путей и соответствующие регуляторные ферменты. 2. Проанализируйте изменения метаболизма, представленные на рис. 11.10 и 11.11 и сравните их с изменениями, изображенными на рис. 11.14. Для этого: а) назовите процессы, которые активируются и ингибируются при длительном голодании; б) выберите и напишите схемы процессов, за счет которых поддерживается концентрация глюкозы в крови при длительном голодании; в) для каждого выбранного процесса укажите ключевые ферменты и гор- моны, под влиянием которых происходит их активация; г) выберите и напишите схемы процессов, за счет которых осуществляет- ся энергообеспечение мышц при длительном голодании. 3. Изучите схему изменений метаболизма при сахарном диабете (рис. 11.16).Объяснитепричинывозникновениягиперглюкоземии.Выпишите названия метаболических путей, которые ускоряются в этих условиях. 4. Объясните причины и механизмы возникновения кетоацидоза при сахарном диабете и изобразите соответствующую схему. 5. Сравните изменения гормонального статуса и метаболизма при сахарном диабете и голодании (рис. 11.14 и 11.16). Объясните, почему на фоне гиперглюкоземии при сахарном диабете происходит катаболизм жиров и белков. 6. Перечислите основные симптомы сахарного диабета. Обоснуйте справедливость выражения: «сахарный диабет - это голод среди изобилия». Для этого: а) назовите проявления сахарного диабета, сходные с изменениями обмена веществ при голодании; б) объясните причины возникновения этих изменений; в) назовите основные различия в обмене веществ при сахарном диабете и голодании. 7. Продолжите заполнение таблицы поздних осложнений сахарного диабета (табл. 11.2): Таблица 11.2. Поздние осложнения сахарного диабета
ЗАДАНИЯ ДЛЯ САМОКОНТРОЛЯ 1. Выберите правильный ответ. При голодании: A. Ацетил-КоА-карбоксилаза фосфорилирована и активна Б. Гормон-чувствительная ТАГ-липаза неактивна B. ЛП-липаза в жировой ткани активна Г. Пируваткиназа в печени фосфорилирована и активна Д. сАМР-зависимая протеинкиназа в адипоцитах активна 2. Выберите правильные ответы. При трехдневном голодании: A. Инсулин-глюкагоновый индекс снижен Б. Скорость глюконеогенеза из аминокислот увеличивается B. Скорость синтеза ТАГ в печени снижается Г. Скорость β-окисления в печени снижается Д. Концентрация кетоновых тел в крови выше нормы 3. Выберите правильные ответы. Увеличение скорости синтеза кетоновых тел при голодании является следствием: A. Снижения уровня глюкагона Б. Снижения образования Ацетил-КоА в печени B. Повышения концентрации жирных кислот в плазме крови Г. Снижения скорости β-окисления в печени Д. Снижения активности гормон-чувствительной ТАГ-липазы в адипоцитах 4. Выберите правильные ответы. При сахарном диабете в печени происходит: A. Ускорение синтеза гликогена Б. Увеличение скорости глюконеогенеза B. Снижение скорости синтеза жиров Г. Увеличение скорости синтеза ацетоацетата Д. Повышение активности ацетил-КоА-карбоксилазы 5. Установите соответствие: A. Высокий уровень инсулина Б. Алкалоз B. Гипоглюкоземия Г. Высокий уровень кортизола Д. Аутоиммунное повреждение β-клеток 1. Только при сахарном диабете 2. Только при голодании 3. Только при стероидном диабете 6. Выберите правильные ответы. При ИЗСД у больных наиболее часто обнаруживается: A. Гиперглюкоземия Б. Высокая скорость катаболизма инсулина B. Концентрация инсулина в крови в норме или выше нормы Г. Антитела к β-клеткам поджелудочной железы Д. Микроангиопатии 7. Установите соответствие: A. Макроангиопатии Б. Катаракта B. Микроангиопатии Г. Нефропатия Д. Нейропатии 1. Активация сорбитольного пути в шванновских клетках 2. Глюкозилирование кристаллинов 3. Утолщение базальных мембран клубочков почек ЭТАЛОНЫ ОТВЕТОВ К «ЗАДАНИЯМ ДЛЯ САМОКОНТРОЛЯ» 1. Д 2. А, Б, В, Д 3. В 4. Б, В, Г 5. 1-Д, 2-В, 3-Г 6. А, Г, Д 7. 1-Д, 2-Б, 3-Г ОСНОВНЫЕ ТЕРМИНЫ И ПОНЯТИЯ 1. Голодание 2. Фазы голодания 3. Сахарный диабет 4. ИЗСД 5. ИНСД 6. Гиперглюкоземия - глюкозурия 7. Кетонемия - кетонурия 8. Азотемия - азотурия 9. Поздние осложнения сахарного диабета 10. Диабетическая кома 11. Кетоацидотическая кома 12. Гиперосмолярная кома 13. Лактоацидотическая кома 14. Микроангиопатии 15. Макроангиопатии 16. Нейропатии 17. Нефропатия ЗАДАНИЯ ДЛЯ АУДИТОРНОЙ РАБОТЫ Решите задачи 1. Туристы не рассчитали запасы продовольствия и пока не добрались до первого населенного пункта вынуждены были голодать в течение 2 суток. Какие изменения в обмене веществ произойдут у этих туристов? Для объяснения: а) укажите, как изменится концентрация глюкозы в крови у туристов к концу 2-го дня голодания; б) напишите схемы процессов, за счет активации которых поддерживается нормальная концентрация глюкозы в первые сутки голодания; в) назовите гормоны, которые регулируют уровень глюкозы в этот период; г) представьте в виде схемы механизм действия этих гормонов; д) укажите регуляторные реакции этих путей и способы их активации. 2. Биохимические исследования крови и мочи больного сахарным диабетом I типа показали:
Как изменятся эти показатели при однократном введении больному средней суточной дозы инсулина? В результате активации каких процессов произойдут эти изменения? 3. На прием к терапевту пришел пациент с жалобами на прогрессирующую слабость, сонливость, головокружение. Симптомы усиливались при голодании, что позволило врачу предположить наличие у больного гипоглюкоземии. Анализ крови подтвердил предположение (уровень глюкозы менее 2,5 ммоль/л) и показал к тому же сильно повышенный уровень С-пептида (более 800 пмоль/л). Пациент не страдает СД и не принимает сахаропонижающих препаратов. Наличие какого заболевания можно предположить? При ответе на вопрос: а) назовите стимулы, влияющие на секрецию инсулина; б) опишите влияние инсулина на углеводный и жировой обмен в печени, жировой ткани и мышцах; в) объясните, чем опасна гипоглюкоземия и какие процессы в организме в норме предотвращают развитие гипоглюкоземии даже при голодании; г) назовите заболевание и предположите метод лечения. 4. Пациент N обратился с жалобами на постоянное чувство голода, жажду, быструю утомляемость и усталость. Определение концентрации глюкозы натощак показало 130 мг/дл. Какие дополнительные исследования для установления диагноза необходимо провести в этом случае? Какие результаты можно прогнозировать в случае обнаружения у обследуемого диабета II типа? 5. Пациент с диагнозом ИЗСД длительное время не получал инъекций инсулина. После обращения больного к врачу и тщательного обследования назначена инсулиновая терапия. Через 2 месяца определение концентрации глюкозы в крови натощак показало 85 мг/дл, уровень глюкозилированного гемоглобина составил 14% общего уровня гемоглобина (норма 5,8-7,2%). Каковы возможные причины высокой концентрации глюкозилированного гемоглобина у данного больного, несмотря на проводимое лечение? Приведите примеры глюкозилирования других белков. Объясните, к каким осложнениям это может привести. 6. Пациент 39 лет поступил с жалобами на сильную жажду, быструю утомляемость. Потеря веса за последние 5 недель составила 4 кг, несмотря на хороший аппетит и нормальную физическую нагрузку. Анализ крови показал, что концентрация глюкозы спустя 2 часа после приема пищи равна 242 мг/дл. Какое заболевание можно предположить у данного пациента? Что является причиной жажды? Как объяснить быструю утомляемость пациента? Модульная единица 3 РЕГУЛЯЦИЯ ВОДНО-СОЛЕВОГО ОБМЕНА. РОЛЬ ВАЗОПРЕССИНА, АЛЬДОСТЕРОНА И РЕНИН-АНГИОТЕНЗИНОВОЙ СИСТЕМЫ. РЕГУЛЯЦИЯ ОБМЕНА СА2+ И ФОСФАТОВ Цели изучения Уметь: 1. Анализировать изменения метаболизма, возникающие при некоторых нарушениях водно-солевого обмена (гиперальдостеронизм, почечная гипертония). 2. Интерпретировать молекулярные механизмы нарушений синтеза и секреции гормонов, обеспечивающих регуляцию обмена кальция. Знать: 1. Характеристики основных гормонов ВСО и этапы его регуляции. 2. Основные функции кальция в организме. 3. Механизмы гормональной регуляции обмена ионов кальция и фосфатов. 4. Проявления некоторых нарушений синтеза и секреции гормонов, регулирующих обмен кальция и фосфатов (гипо- и гиперпаратиреоидизм, рахит). ТЕМА 11.8. РЕГУЛЯЦИЯ ВОДНО-СОЛЕВОГО ОБМЕНА 1. Основными параметрами водно-солевого гомеостаза являются осмотическое давление, рН и объем внутриклеточной и внеклеточной жидкости. Изменение этих параметров может привести к изменению кровяного давления, ацидозу или алкалозу, дегидратации и отекам. Основными гормонами, участвующими в регуляции водно-солевого баланса, являются антидиуретический гормон (АДГ), альдостерон и предсердный натриуретический фактор (ПНФ). 2. Антидиуретический гормон (АДГ), или вазопрессин, - пептид, содержащий девять аминокислот, соединенных одним дисульфидным мостиком. Синтезируется в виде прогормона в гипоталамусе, затем переносится в нервные окончания задней доли гипофиза, из которых секретируется в кровоток при соответствующей стимуляции. Перемещение по аксону связано со специфическим белком-переносчиком (нейрофизином) (рис. 11.17). Стимулом, вызывающим секрецию АДГ, служит повышение концентрации ионов натрия и увеличение осмотического давления внеклеточной жидкости. Наиболее важные клетки-мишени для АДГ - клетки дистальных канальцев и собирательные трубочки почек. Клетки этих протоков относительно непроницаемы для воды, и в отсутствие АДГ моча не концентрируется и может выделяться в количествах, превышающих 20 л в сутки (норма 1-1,5 л в сутки).
А: 1 - супраоптический нейрон; 2 - паравентрикулярный нейрон; 3 - передняя доля гипофиза; 4 - задняя доля гипофиза; 5 - АДГ-нейрофизин; Б: 1 - АДГ связывается с мембранным рецептором V2, вызывая активацию аденилатциклазы (АЦ) и, как следствие, образование цАМФ; 2 - цАМФ активирует протеинкиназу, фосфорилирующую белки; 3 - фосфорилированные белки индуцируют транскрипцию гена белка аквапорина; 4 - аквапорин встраивается в мембрану клетки почечного канальца Для АДГ существует два типа рецепторов - V1 и V2. Рецептор V2обнаружен только на поверхности эпителиальных клеток почек. Связывание АДГ с V2 сопряжено с аденилатциклазной системой и стимулирует активацию протеинкиназы (ПКА), которая фосфорилирует белки, стимулирующие экспрессию гена мембранного белка - аквапорина-2. Аквапорин-2 перемещается к апикальной мембране, встраивается в нее и образует водные каналы, через которые молекулы воды свободно диффундируют в клетки почечных канальцев, а затем поступают в интерстициальное пространство. В результате происходит реабсорбция воды из почечных канальцев (см. рис. 11.17). Рецепторы типа V локализованы в мембранах гладких мышц. Взаимодействие АДГ с рецептором V1 приводит к активации фосфолипазы С, в результате чего происходит высвобождение Са2+ из эндоплазматического ретикулума и сокращение гладкомышечного слоя сосудов. 3. Несахарный диабет. Дефицит АДГ, вызванный дисфункцией задней доли гипофиза, а также нарушением в системе передачи гормонального сигнала, может приводить к развитию несахарного диабета. Основным проявлением несахарного диабета является полиурия, т.е. выделение большого количества мочи низкой плотности. 4. Альдостерон - наиболее активный минералокортикостероид - синтезируется клетками клубочковой зоны коры надпочечников из холестерола. Синтез и секреция альдостерона стимулируется при низкой концентрации Na+, высокой концентрации К+ и ренин-ангиотензиновой системой. Гормон проникает внутрь клеток почечных канальцев, взаимодействует со специфическим рецептором, цитоплазматическим или ядерным (рис. 11.18), и индуцирует синтез белков, которые обеспечивают реабсорбцию ионов натрия и экскрецию ионов калия. Кроме того, белки, синтез которых индуцируется альдостероном, увеличивают количество насосов Na+, K+ - АТФазы, а также служат ферментами ЦТК, генерирующего молекулы АТФ для активного транспорта ионов. Суммарным результатом действия альдостерона является задержка NaCl в организме. 5. Главную роль в регуляции водно-солевого баланса, а значит, регуляции объема крови и артериального давления служит система ренин-ангиотензинальдостерон (рис. 11.19). Протеолитический фермент ренин синтезируется юкстагломерулярными клетками почечных афферентных артериол. Уменьшение кровяного давления в приносящих артериолах, потеря жидкости или крови, снижение концентрации NaCl стимулируют высвобождение ренина. Образующийся в печени белок ангиотензиноген гидролизуется ренином с образованием ангиотензина I, который в свою очередь служит субстратом для АПФ (ангиотензин-превращающего фермента карбоксидипентидилпептидазы). От ангиотензина I отщепляется дипептид с образованием ангиотензина II. Через инозитолфосфатную систему ангиотензин II стимулирует синтез и секрецию альдостерона. Являясь также мощным сосудосуживающим веществом, ангиотензин II вызывает сокращение гладкомышечных клеток кровеносных сосудов, соответственно повышение кровяного давления и, кроме этого, вызывает жажду. 6. Система ренин-ангиотензин-альдостерон обеспечивает восстановление объема крови, который может уменьшиться в результате кровотечения, обильной рвоты, диареи, потения - состояний, являющихся сигналом для
Альдостерон, взаимодействуя с внутриклеточными рецепторами, стимулирует синтез белков. Эти белки могут быть: 1 - компонентами натриевых каналов и увеличивать реабсорбцию Na+ из мочи; 2 - ферментами ЦТК, активность которых обеспечивает продукцию АТР; 3 - Na+, K+ - АТФазой, насосом, который поддерживает низкую внутриклеточную концентрацию ионов натрия и высокую концентрацию ионов калия высвобождения ренина. Этому способствует также снижение импульсации от барорецепторов предсердий и артерий в результате уменьшения внутрисосудистого объема жидкости. В результате увеличивается образование ангиотензина II и соответственно повышается в крови концентрация альдостерона, вызывая задержку ионов натрия. Это служит сигналом для осморецепторов гипоталамуса и секреции из нервных окончаний передней доли гипофиза АДГ, который стимулирует реабсорбцию воды из собирательных трубочек. Ангиотензин II, оказывая сильное сосудосуживающее действие, повышает артериальное давление, а также усиливает жажду. Поступающая с питьем вода в большей мере, чем это происходит в норме, задерживается в организме.
АПФ - ангиотензинпревращающий фермент (другое название карбоксипептидилдипептидаза) • уменьшение объема жидкости и снижение артериального давления активируют систему ренин-ангиотензин-альдостерон; • ангиотензин II вызывает кратковременное сужение сосудов и повышение кровяного давления; • альдостерон стимулирует задержку натрия, вследствие чего происходит высвобождение вазопрессина и усиливается реабсорбция воды; • ангиотензин II вызывает также чувство жажды, что способствует увеличению жидкости в организме Увеличение объема жидкости и повышение кровяного давления приводит к устранению стимула, который вызвал активацию ренин-ангиотензиновой системы и секрецию альдостерона и, как результат, приводит к восстановлению объема крови. 7. Снижение перфузионного давления в почечных клубочках может наступить и вследствие сужения (стеноза) почечной артерии или нефросклероза. В этом случае также включается вся ренин-ангиотензиновая система. Но поскольку исходные объем и давление крови при этом нормальные, включение системы приводит к повышению кровяного давления сверх нормы и развитию так называемой почечной гипертонии. 8. Гиперальдостеронизм - это заболевание, вызванное гиперсекрецией альдостерона надпочечниками. Причиной первичного гиперальдостеронизма (синдром Кона) является аденома надпочечников или диффузная гипертрофия клеток клубочковой зоны, вырабатывающих альдостерон. При первичном гиперальдостеронизме избыток альдостерона усиливает реабсорбцию натрия в почечных канальцах. Повышение концентрации Na+ в плазме служит стимулом к секреции антидиуретического гормона и задержке воды почками. Кроме того, усиливается выведение ионов калия, магния и протонов. В результате развивается гипернатриемия, вызывающая, в частности, гипертонию, гиперволемию и отеки; гипокалиемия, ведущая к мышечной слабости, а также дефицит магния и метаболический алкалоз. Причиной вторичного гиперальдостеронизма является повышенный уровень ренина и анигиотензина II, это стимулирует кору надпочечников и приводит к избыточному синтезу альдостерона. Клинические симптомы менее выражены, чем при первичном альдостеронизме. Одновременное определение концентрации альдостерона и активности ренина в плазме позволяет окончательно дифференцировать первичный (активность ренина в плазме снижена) и вторичный (активность ренина в плазме повышена) гиперальдостеронизм. 9. Предсердный натриуретический фактор (ПНФ) - пептид, который синтезируется и хранится в виде прогормона в кардиоцитах. Основной фактор, регулирующий секрецию ПНФ, - увеличение артериального давления. Основные клетки-мишени ПНФ - почки, надпочечники, периферические артерии. Рецептор ПНФ плазматической мембраны является каталитическим рецептором, обладающим активностью гуанилатциклазы. В результате
1 - ингибирует выделение ренина; 2 - ингибирует секрецию альдостерона; 3 - ингибирует секрецию АДГ; 4 - вызывает релаксацию сосудов связывания ПНФ с рецептором гуанилатциклазная активность рецептора возрастает и происходит образование из ГТФ циклического ГМФ. В результате действия ПНФ ингибируется образование и секреция ренина и альдостерона. Суммарным эффектом действия ПНФ является увеличение экскреции Na+ и воды и понижение кровяного давления (рис. 11.20). ПНФ обычно рассматривают как физиологический антагонист ангиотензина II, так как он вызывает расширение сосудов и потерю соли и воды. ТЕМА 11.9. РЕГУЛЯЦИЯ ОБМЕНА КАЛЬЦИЯ И ФОСФАТОВ. СТРОЕНИЕ, СИНТЕЗ И МЕХАНИЗМ ДЕЙСТВИЯ ПАРАТГОРМОНА, КАЛЬЦИТРИОЛА И КАЛЬЦИТОНИНА 1. В организме взрослого человека содержится -1,2 кг кальция. Основной фонд кальция в организме - кальций костей (99% всего кальция в организме). Другой фонд - ионы кальция, растворенные в жидкостях или соединенные с белками жидкостей и тканей. Концентрация кальция внутри клеток зависит от его концентрации во внеклеточной жидкости. Концентрация Са2+ в крови здоровых людей составляет 2,12-2,6 ммоль/л (9-11 мг/дл), во внутриклеточной жидкости - в тысячи раз меньше. Кальций служит основным минеральным структурным компонентом костной ткани. Ионы кальция участвуют в мышечном сокращении, повышают проницаемость мембраны клеток для ионов калия, влияют на натриевую проводимость клеток, на работу ионных насосов, способствуют секреции гормонов, участвуют в каскадном механизме свертывания крови, служат важнейшими посредниками во внутриклеточной передаче сигналов. Концентрация Са2+ в плазме регулируется с высокой точностью: изменение ее всего на 1% приводит в действие гомеостатические механизмы, восстанавливающие равновесие. Основными регуляторами обмена Са2+ в крови являются паратгормон, кальцитриол и кальцитонин. 2. Паратгормон синтезируется паращитовидными железами в виде препрогормона, который затем превращается в зрелый гормон путем частичного протеолиза. ПТГ секретируется в ответ на снижение концентрации кальция в крови. Основными органами-мишенями для гормона являются кости и почки (рис. 11.21). Гормон инициирует каскад событий, связанный с аденилатциклазой остеобластов, которые стимулируют метаболическую активность остеокластов. Происходит мобилизация Са2+ из кости и поступление фосфатов в кровь, а в дистальных канальцах почек стимулируется реабсорбция Са2+ и уменьшается реабсорбция фосфатов, в результате чего восстанавливается нормальный уровень ионов кальция во внеклеточной жидкости. 3. Кальцитриол, как и другие стероидные гормоны, синтезируется из холестерола. Непосредственным предшественником кальциферола является холекальциферол (витамин D3). Небольшое количество витамина D3 содержится
1 - ПТГ стимулирует мобилизацию кальция из кости; 2 - ПТГ стимулирует реабсорбцию ионов кальция в дистальных канальцах почек; 3 - ПТГ активирует образование 1,25(OH)2D3 в почках, что приводит к стимуляции всасывания Са2+ в кишечнике в продуктах питания, но большая часть витамина, используемого в синтезе кальцитриола, образуется в коже из 7-дегидрохолестерола в ходе неферментативной реакции под действием ультрафиолетового света. Образование кальцитриола из витамина D3 начинается в печени и заканчивается в почках (рис. 11.22). В печени холекальциферол гидроксилируется по 25-му атому углерода с образованием 25-гидроксихолекальциферола. Гидроксилирование, протекающее в почках под действием фермента 1о-гидроксилазы, является скорость лимитирующей стадией и приводит к образованию кальцитриола 1,25(OH)2D3 - активной формы витамина D3. Фермент этой реакции активируется низкой концентрацией в крови ионов Са2+ и паратгормоном. Увеличение концентрации кальцитриола, напротив, тормозит синтез 1о-гидроксилазы почек, ингибируя образование гормона. Транспортируясь по крови в комплексе с белком-переносчиком, кальцитриол связывается с внутриклеточным рецептором, взаимодействует с хроматином и изменяет скорость трансляции. В результате в клетках-мишенях синтезируются белки, обеспечивающие всасывание кальция и фосфатов в энтероциты. 4. Кальцитонин - полипептид, состоящий из 32 аминокислотных остатков с одной дисульфидной связью. Гормон секретируется парафолликулярными
1 - холестерол является предшественником кальцитриола; 2 - в коже 7-дегидрохолестерол неферментативно под действием УФ-облучения превращается в холекальциферол; 3 - в печени 25-гидроксилаза превращает холекальциферол в кальцидиол; 4 - в почках образование кальцитриола катализируется 1о-гидроксилазой К-клетками щитовидной железы или С-клетками паращитовидной железы в виде высокомолекулярного белка-предшественника. Секреция кальцитонина возрастает при увеличении концентрации Са2+ и уменьшается при понижении концентрации Са2+ в крови. Кальцитонин ингибирует высвобождение Са2+ из кости и стимулирует его экскрецию почками с мочой. 5. Гипокальциемия и гиперкальциемия, когда концентрация кальция в плазме крови ниже или выше нормы, свидетельствует о патологии. Изменение уровня кальция в крови влияет на концентрацию кальция внутри клеток, что приводит к изменению порога возбудимости нервных и мышечных клеток, нарушению функционирования кальциевого насоса, снижению активности ферментов и нарушению гормональной регуляции метаболизма. При гипокальциемии наблюдаются гиперрефлексы, судороги, спазмы гортани. При гиперкальциемии наблюдается снижение нервно-мышечной возбудимости, может наступить глубокое расстройство нервных функций, психозы, ступор, кома. 6. Гиперпаратиреоз. Избыточная секреция паратгормона, возникающая в результате опухоли околощитовидной железы, диффузной гиперплазии желез, карциномы паращитовидной железы (первичный гиперпаратиреоз), приводит к повышению мобилизации кальция и фосфатов из кости, усилению реабсорбции кальция и выведению фосфатов в почках. Вследствие этого возникает гиперкальциемия, которая может приводить к снижению нервно-мышечной возбудимости и мышечной гипотонии. У больных появляется общая и мышечная слабость, быстрая утомляемость и боли в отдельных группах мышц, увеличивается риск переломов позвоночника, бедренных костей и костей предплечья. Увеличение концентрации фосфата и ионов кальция в почечных канальцах может служить причиной образования в почках камней и приводит к гиперфосфатурии и гипофосфатемии. 7. Гипопаратиреоз. Основным симптомом гипопаратироза, обусловленного недостаточностью паращитовидных желез, является гипокальциемия. Понижение концентрации ионов кальция в крови может вызвать неврологические, офтальмологические и сердечно-сосудистые нарушения, а также поражения соединительной ткани. У больного гипопаратиреозом отмечается повышение нервно-мышечной проводимости, приступы тонических судорог, судороги дыхательных мышц и диафрагмы, ларингоспазм. 8. Рахит - заболевание детского возраста, связанное с недостаточной минерализацией костной ткани. Нарушение минерализации кости является следствием дефицита кальция и может быть обусловлено следующими причинами: недостатком витамина D3 в пищевом рационе, нарушением всасывания витамина D3 в тонком кишечнике, снижением синтеза предшественников кальцитриола из-за недостаточного времени пребывания на солнце, дефектом Ια-гидроксилазы, дефектом рецепторов кальцитриола в клетках-мишенях. Все это вызывает снижение всасывания кальция в кишечнике и снижение его концентрации в крови, стимуляцию секреции паратгормона и вследствие этого - мобилизацию ионов кальция из кости. При рахите поражаются кости черепа, грудная клетка вместе с грудиной выступает вперед, деформируются трубчатые кости и суставы рук и ног, увеличивается и выпячивается живот. Основным способом предупреждения рахита является правильное питание и достаточная инсоляция. ЗАДАНИЯ ДЛЯ ВНЕАУДИТОРНОЙ РАБОТЫ Решите задачи 1. Изучите механизмы регуляция поддержания водного баланса, запомните стимулы, вызывающие секрецию гормонов и особенности механизма их действия (рис. 11.19). Изобразите в виде схемы последовательность событий при восстановлении водно-солевого равновесия после приема соленой пищи. 2. У мужчины 23 лет при проведении хирургической операции по удалению опухоли из верхнего отдела передней доли гипофиза был затронут перешеек задней доли гипофиза. В послеоперационный период у пациента развилась полиурия. Как можно объяснить появление этого симптома у данного пациента? Для обоснования ответа: а) назовите гормоны, синтезируемые в гипоталамусе и секретируемые из задней доли гипофиза; б) нарисуйте схему передачи сигнала этого гормона на клетки-мишени; в) назовите эффекты этого гормона. 3. Вспомните схему синтеза стероидных гормонов (рис. 11.8) и выпишите в тетрадь последовательность этапов синтеза альдостерона. 4. Составьте свою схему, иллюстрирующую эффекты альдостерона и механизм его действия. 5. Изучите схему регуляции синтеза и секреции альдостерона с участием системы ренин-ангиотензин (рис. 11.19) и подберите недостающие компоненты, обозначенные на схеме (рис. 11.23) цифрами. 6. Составьте свою схему, объясняющую основные результаты действия ПНФ (рис. 11.20) и ответьте на вопрос, на чем основан гипотензивный эффект ПНФ. 7. Заполните табл. 11.3. Таблица 11.3. Характеристика гормонов, регулирующих водно-солевой обмен
8. Заполните табл. 11.4. Таблица 11.4. Характеристика гормонов, регулирующих обмен кальция и фосфатов
9. Используя схему рис. 11.22, укажите все возможные причины рахита и представьте схему механизма передачи сигнала кальцитриола на клеткимишени. 10. При гиповитаминозе D3 нарушается процесс минерализации костей, уменьшается содержание в них кальция и фосфатов; концентрация Са2+ в крови сохраняется в пределах нормы или несколько снижается. Составьте схему поддержания гомеостаза Са2+ при гиповитаминозе D3 и определите: а) за счет каких источников поддерживается нормальная концентрация Са2+ в крови в этом случае; б) как изменится концентрация в крови кальцитонина и паратгормона. 11. Увеличение выведения кальция с мочой может быть причиной образования почечных камней, состоящих преимущественно из оксалата кальция. Назовите причины, по которым может увеличиваться выведение Са2. ЗАДАНИЯ ДЛЯ САМОКОНТРОЛЯ 1. Выберите правильный ответ. В ответ на повышение осмотического давления возрастает синтез и секреция гормона: A. Альдостерона Б. Кортизола B. Вазопрессина Г. Адреналина Д. Глюкагона 2. Установите соответствие. Место синтеза: A. Печень Б. Почки B. Гипоталамус Г. Надпочечники Д. Поджелудочная железа Метаболиты: 1. Вазопрессин 2. Альдостерон 3. Ренин 3. Установите соответствие: A. Стимул для синтеза и секреции - образование ангиотензина II Б. Стимул для секреции - повышение концентрации ионов натрия B. Органы-мишени - периферические артерии Г. Гиперпродукция гормона приводит к полиурии Д. Место синтеза - печень 1. Вазопрессин 2. Альдостерон 3. Ангиотензиноген 4. Выберите правильные ответы. Ангиотензин II: A. Образуется в печени Б. Является протеолитическим ферментом B. Является субстратом ренина Г. Стимулирует синтез альдостерона Д. Стимулирует сужение сосудов 5. Выберите правильные ответы. Кальцитриол: A. Стимулирует реабсорбцию кальция в почках Б. Является предшественником 7-дегидрохолестерола B. Стимулирует реабсорбцию натрия в почках Г. Увеличивает скорость всасывания кальция в кишечнике Д. Стимулирует мобилизацию кальция из костей 6. Выберите правильные ответы. Снижение концентрации Са2+ в плазме крови вызывает: A. Увеличение секреции паратгормона Б. Ингибирование активности парафолликулярных клеток щитовидной железы B. Гидроксилирование метаболитов витамина D3 Г. Уменьшение экскреции кальция почками Д. Повышение скорости резорбции кости 7. Выполните «цепное» задание: а) в гипоталамусе синтезируется гормон: A. Вазопрессин Б. Адреналин B. Альдостерон Г. Кальцитриол б) клетками-мишенями для данного гормона являются: A. Клетки ЮГА Б. Периферические артерии B. Клетки собирательных трубочек и дистальных канальцев Г. Клетки клубочка нефрона в) связываясь с рецепторами этих клеток, он стимулирует: A. Аденилатциклазную систему Б. Фосфопротеинфосфатазу B. Инозитолтрифосфатную систему Г. Ренин-ангиотензиновую систему. г) в результате активации этой системы увеличивается количество белка: A. Альбумина Б. Транспортеров натрия B. Аквапорина-2 Г. Транспортера калия д) этот белок обеспечивает увеличение реабсорбции: A. Ионов калия Б. Ионов кальция B. Ионов натрия Г. Воды 8. Выберите правильные ответы. Паратгормон: A. Транспортируется по крови в комплексе с белком-переносчиком Б. Секреция регулируется концентрацией кальция в крови B. Недостаточность гормона приводит к снижению концентрации Са2+ Г. Для проявления биологической активности необходима вся молекула гормона целиком Д. Увеличивает эффективность всасывания воды в кишечнике 9. Выберите правильные ответы. Вазопрессин: A. Стимулирует повышение осмотического давления плазмы крови Б. Активирует протеинкиназу С в почках B. Стимулирует реабсорбцию воды в почках Г. Снижает осмотическое давление плазмы крови Д. Стимулирует экспрессию гена аквапорина-2 10. Установите соответствие: A. Проявляет сосудосуживающий эффект Б. Стимулирует реабсорбцию Na+ B. Взаимодействует с мембранными рецепторами клеток-мишеней Г. Усиливает секрецию ренина Д. Является протеолитическим ферментом 1. Альдостерон 2. Ангиотензин II 3. Ренин 11. Выберите все правильные ответы. ПНФ: A. Взаимодействует с мембранными рецепторами клеток-мишеней Б. Активирует фосфолипазу С B. Активирует гуанилатциклазу Г. Подавляет секрецию альдостерона Д. Увеличивает выведение воды и Na+ 12. Установите соответствие: A. В почках Б. В коже B. В печени Г. В мозге Д. В кишечнике 1. Превращение 7-дегидрохолестерина в витамин D3 путем неферментативного фотолиза 2. Образование 1,25 (ОН)2D3 в монооксигеназной реакции с участием NADPH 3. Индукция синтеза кальций-связывающего белка ЭТАЛОНЫ ОТВЕТОВ К «ЗАДАНИЯМ ДЛЯ САМОКОНТРОЛЯ» 1. В 7. а) А, б) В, в) А, г) В, д) Г 2. 1-В; 2-Г; 3-Б 8. Б, В 3. 1-Б; 2-А; 3-Д 9. В, Г, Д 4. Г, Д 10. 1-Б; 2-А; 3-Д 5. А, Г, Д 11. А, В, Г, Д 6. А, В, Г, Д 12.1 - Б; 2 - В; 3 - Д ОСНОВНЫЕ ТЕРМИНЫ И ПОНЯТИЯ 1. Водно-солевой гомеостаз 2. Несахарный диабет 3. Система ренин-ангиотензин-альдостерон 4. Гиперальдостеронизм 5. Гиперкальциемия 6. Гипокальциемия 7. Гипопаратиреоз 8. Гиперпаратиреоз 9. Рахит 10. ПНФ 11. АПФ ЗАДАНИЯ ДЛЯ АУДИТОРНОЙ РАБОТЫ Решите задачи 1. Некоторые формы гипертонии возникают вследствие различных почечных нарушений, например при сдавлении опухолью почечной артерии. Основным методом лечения в подобных случаях является удаление пораженного органа (почки). Однако улучшение состояния больных отмечается при назначении больным препаратов, являющихся ингибиторами АПФ. Нарисуйте схему, отражающую изменение водно-солевого обмена при сдавлении почечной артерии. В результате каких изменений наступает улучшение состояния больных? 2. К врачу обратился пациент с жалобами на частое мочеиспускание и постоянное чувство жажды. При обследовании отмечено увеличение суточного объема мочи при резком снижении ее плотности. Проведенный анализ показал, что уровень инсулина в пределах нормы, но выявлено повышение содержание гормона, ответственного за реабсорбцию воды. Предположите причину полиурии у данного больного? Для ответа на вопрос: а) назовите этот гормон; б) перечислите стимулы, вызывающие его секрецию; в) назовите типы рецепторов для этого гормона и места их локализации; г) приведите схему передачи сигнала данного гормона в почках; д) опишите эффекты гормона в тканях-мишенях; е) приведите схему регуляции секреции этого гормона. 3. Мужчина 48 лет обратился к врачу с жалобами на слабость, мышечные боли, запоры и недавно появившиеся приступы болей в спине и при мочеиспускании. При обследовании больному поставлен диагноз: первичный гиперпаратиреоидизм как следствие развития гиперсекреторной доброкачественной опухоли левой доли паращитовидной железы. Объясните, почему при гиперпаратироидизме может развиться почечнокаменная болезнь? При решении задачи используйте схемы к заданию 5. 4. К педиатру обратилась женщина с жалобами на то, что ее двухлетний сын стал капризным, раздражительным, плохо ест. Появилась потливость, стул неустойчивый. При осмотре установлена податливость костей черепа, деформация грудной клетки. В биохимическом анализе крови уровень общего кальция - 1,57 ммоль/л (норма 2,3-2,8 ммоль/л). Предположите, каким заболеванием страдает этот ребенок. Для этого: а) сравните количество общего кальция в крови у ребенка с нормой, дайте название этому состоянию; б) укажите возможные причины, которые могут привести к развитию данного заболевания; в) приведите схему синтеза гормональной регуляции обмена кальция; г) укажите механизм действия гормонов, причины и последствия их недостаточности в организме; д) дайте рекомендации для дальнейшего обследования и лечения данного заболевания ребенка. 5. Изучите схему: Причины и последствия гипопаратиреоидизма (рис. 11.24). Составьте аналогичные схемы для: а) гиперпаратиреоидизма; б) рахита
Рис. 11.24. Причины и последствия гипопаратиреоидизма Источник: Ваша оценка:
|
Пользователей онлайн: 0. Всего гостей: 0 Поисковики: нет. 
Приветствую вас на своем сайте, здесь вы можете найти много полезной информации (или что-то типа того) САЗОНОВ Вячеслав Фёдорович доцент кафедры биологии Рязанского государственного университета имени С.А. Есенина, кандидат биологических наук. Преподаватель вуза с 1978 года... РекламаПоискПритча наудачу:На сайте введена регистрация через социальные сети, если вы хотите оставлять комментарии без потверждения, пожалуйста, воспользуйтесь именно этим типом аутентификации. Если у вас уже есть аккаунт на сайте, вы можете привязать его к любой социальной сети? зайдя в настройки вашего аккаунта("Мои учётные данные") ниже и воспользовавшись вкладкой "Подключение к социальным сетям". После того, как вы зайдёте при помощи аккаунта в социальной сети, ваши возможности на сайте возрастут. Поддержка сайтаВы можете поддержать сайт не только добрым словом, но и материально! Это очень поможет. IT-специалисты, следящие за сайтом день и ночь, хотя бы лишнюю чашечку кофе выпьют. Для этого по своему желанию перечислите любую сумму на карту Сбербанка номер: 2202 2008 3795 8501 -24-04-26-03-08 |
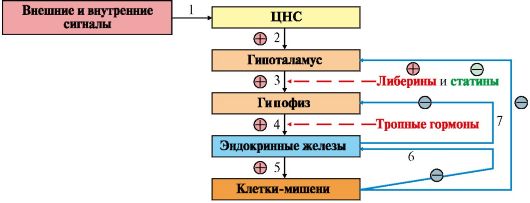
 Изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной обратной связи подавляет синтез гормонов, действуя либо на эндокринные железы, либо на гипоталамус; синтез и секреция тропных гормонов подавляется гормонами периферических желез.
Изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной обратной связи подавляет синтез гормонов, действуя либо на эндокринные железы, либо на гипоталамус; синтез и секреция тропных гормонов подавляется гормонами периферических желез.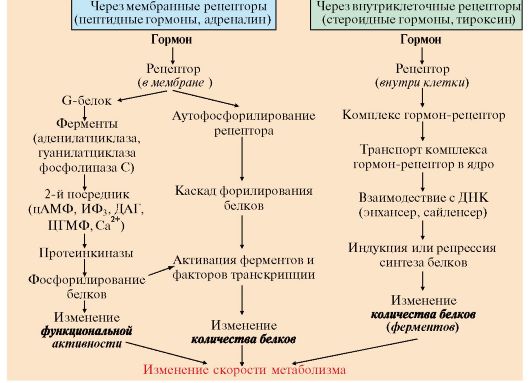 Рис. 11.2. Основные этапы передачи гормональных сигналов в клетки-мишени
Рис. 11.2. Основные этапы передачи гормональных сигналов в клетки-мишени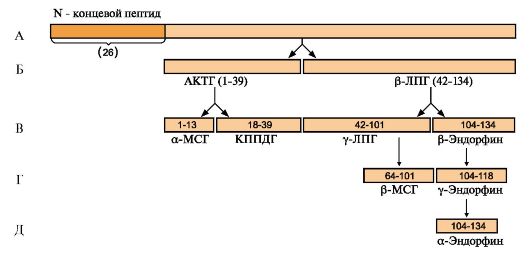 Рис. 11.3. Образование пептидных гормонов, являющихся продуктами общего гена:
Рис. 11.3. Образование пептидных гормонов, являющихся продуктами общего гена: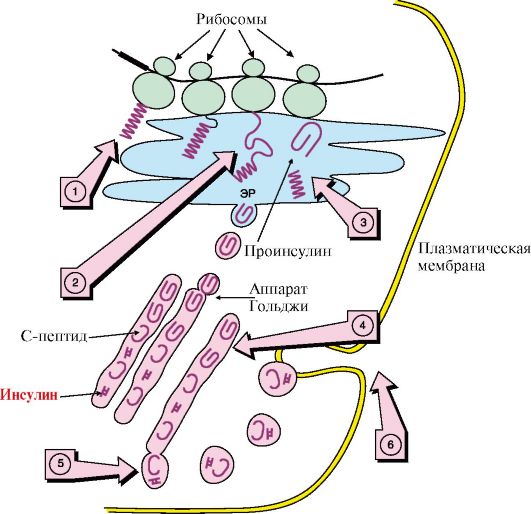 Рис. 11.4. Схема биосинтеза инсулина в клетках поджелудочной железы:
Рис. 11.4. Схема биосинтеза инсулина в клетках поджелудочной железы: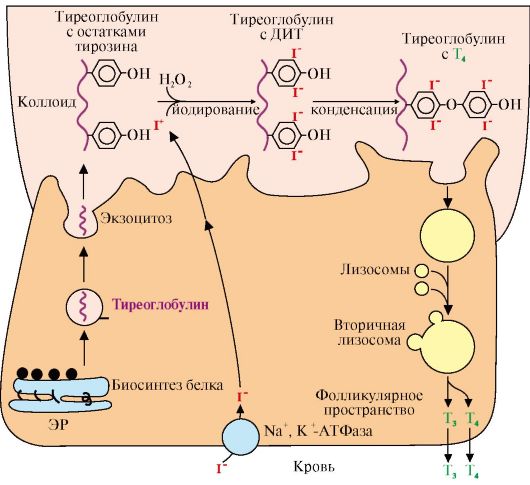 Рис. 11.5. Синтез йодтиронинов:
Рис. 11.5. Синтез йодтиронинов: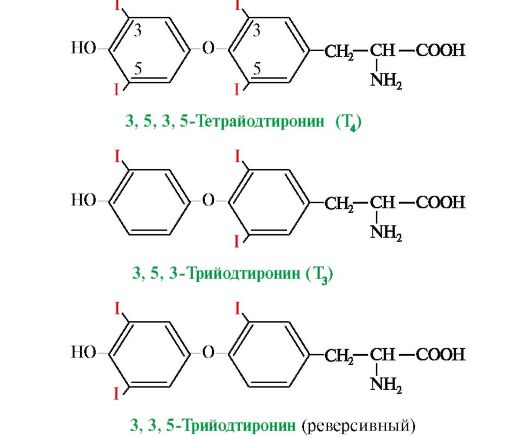 Рис. 11.6. Структура гормонов щитовидной железы
Рис. 11.6. Структура гормонов щитовидной железы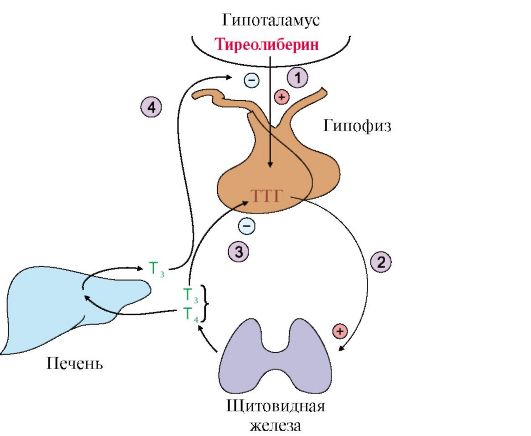 Рис. 11.7. Регуляция синтеза и секреции йодтиронинов:
Рис. 11.7. Регуляция синтеза и секреции йодтиронинов: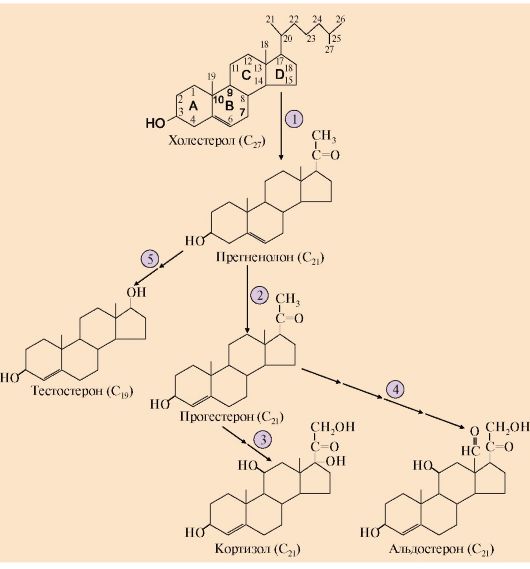 Рис. 11.8. Синтез основных кортикостероидов:
Рис. 11.8. Синтез основных кортикостероидов: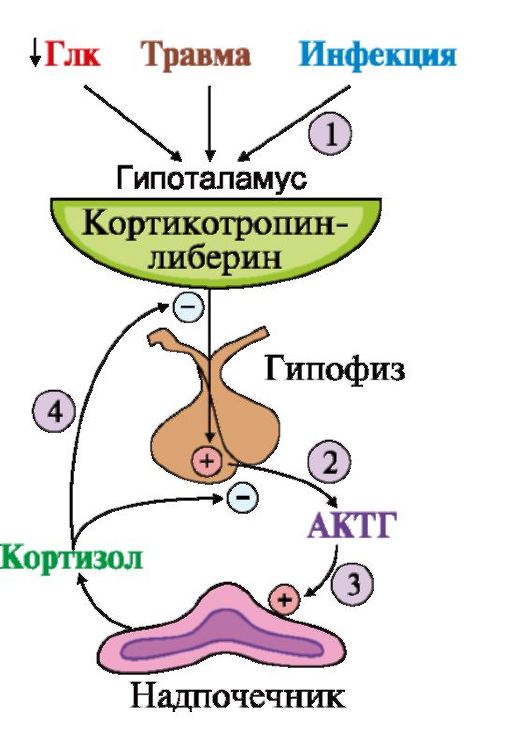 Рис. 11.9. Регуляция синтеза и секреции кортизола:
Рис. 11.9. Регуляция синтеза и секреции кортизола: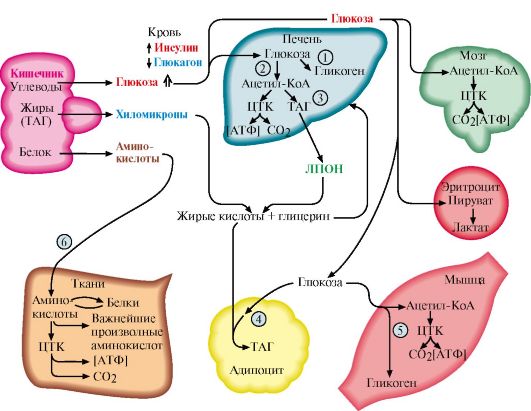 Рис. 11.10. Пути использования основных энергоносителей в абсорбтивном периоде:
Рис. 11.10. Пути использования основных энергоносителей в абсорбтивном периоде: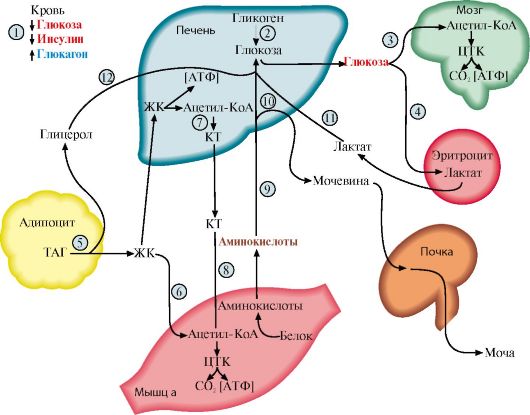 Рис. 11.11. Пути использования основных энергоносителей при смене абсорбтивного состояния на постабсорбтивное:
Рис. 11.11. Пути использования основных энергоносителей при смене абсорбтивного состояния на постабсорбтивное: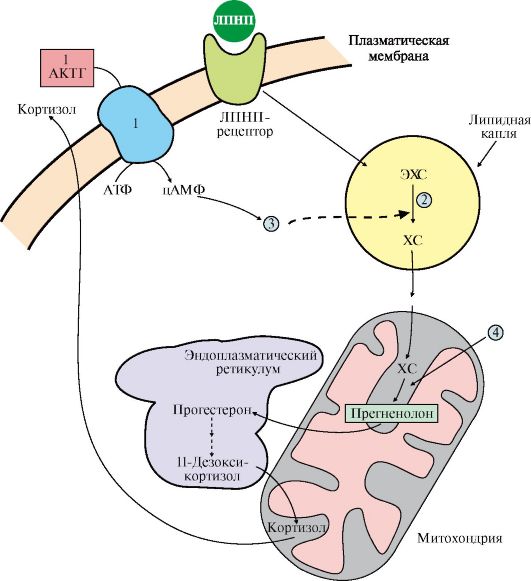 Рис. 11.12. Внутриклеточный цикл синтеза кортизола:
Рис. 11.12. Внутриклеточный цикл синтеза кортизола: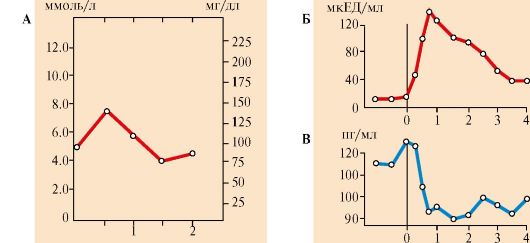 Рис. 11.13. Динамика изменений концентрации глюкозы (А), инсулина (Б) и глюкагона (В) после приема пищи, богатой углеводами
Рис. 11.13. Динамика изменений концентрации глюкозы (А), инсулина (Б) и глюкагона (В) после приема пищи, богатой углеводами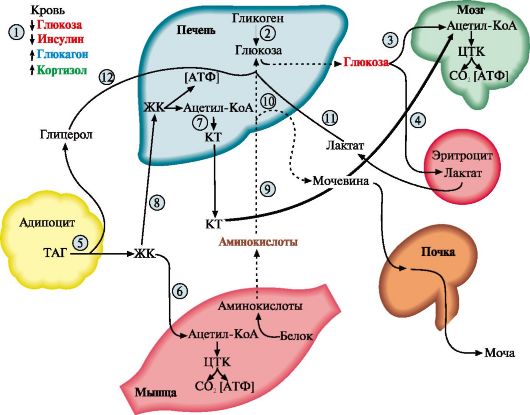 Рис. 11.14. Изменения метаболизма основных энергоносителей при голодании:
Рис. 11.14. Изменения метаболизма основных энергоносителей при голодании: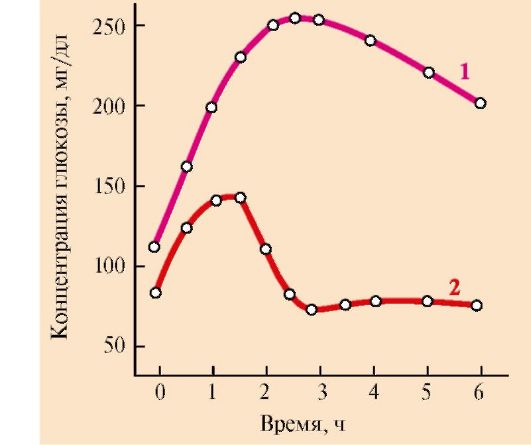 Рис. 11.15. Изменение толерантности к глюкозе у больных скрытой формой сахарного диабета.
Рис. 11.15. Изменение толерантности к глюкозе у больных скрытой формой сахарного диабета.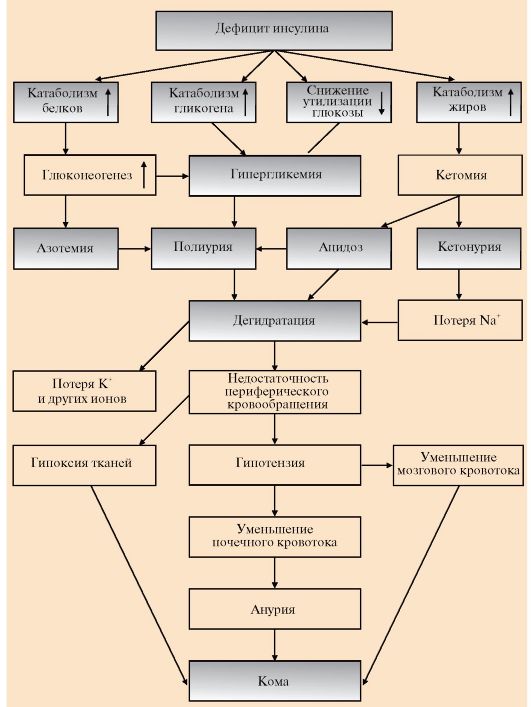 Рис. 11.16. Изменения метаболизма при сахарном диабете и причины диабетической комы
Рис. 11.16. Изменения метаболизма при сахарном диабете и причины диабетической комы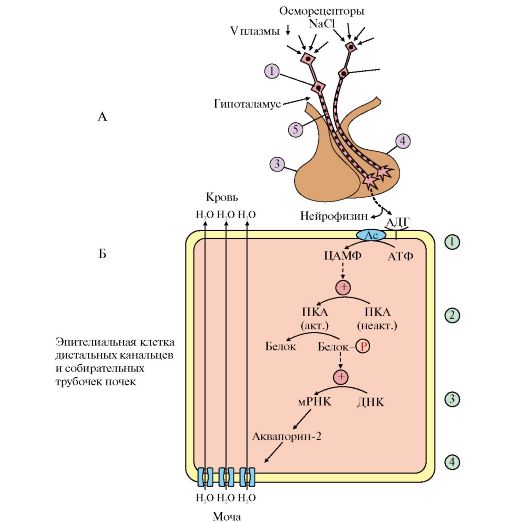 Рис. 11.17. Секреция и механизм действия антидиуретического гормона:
Рис. 11.17. Секреция и механизм действия антидиуретического гормона: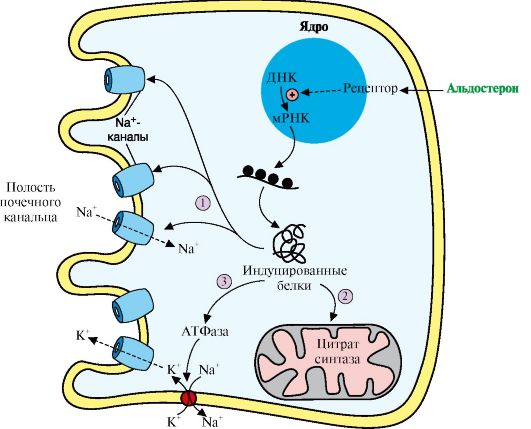 Рис. 11.18. Механизм действия альдостерона.
Рис. 11.18. Механизм действия альдостерона.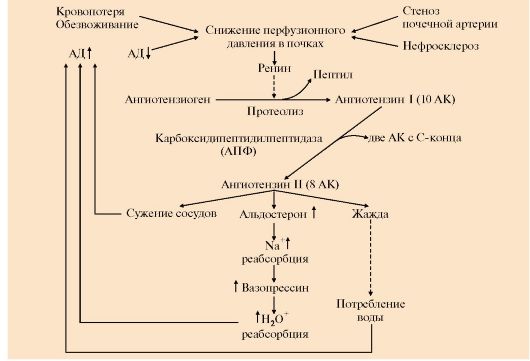 Рис. 11.19. Система ренин-ангиотензин-альдостерон.
Рис. 11.19. Система ренин-ангиотензин-альдостерон.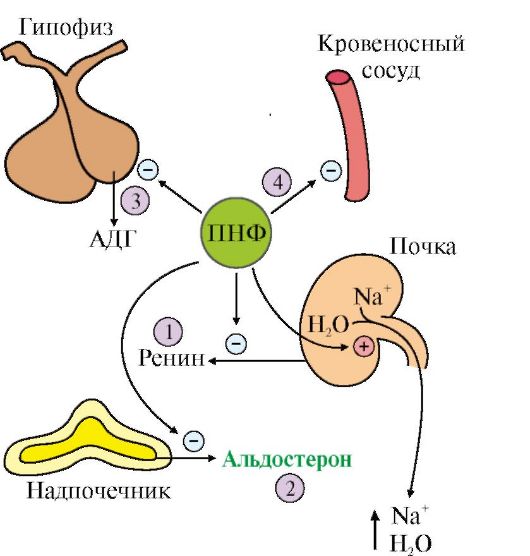 Рис. 11.20. Эффекты действия ПНФ:
Рис. 11.20. Эффекты действия ПНФ: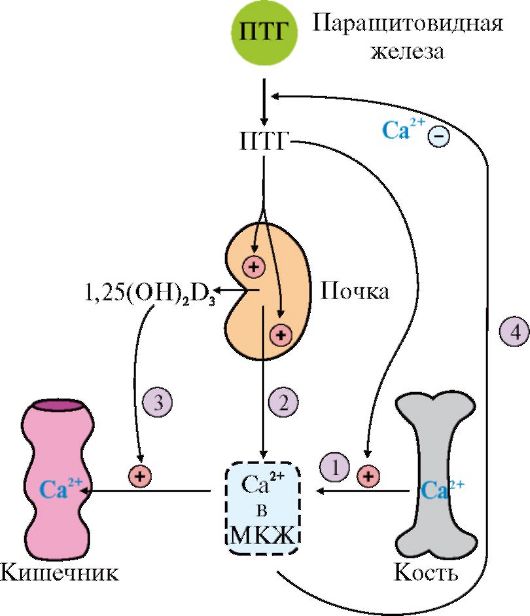 Рис. 11.21 Эффекты действия ПТГ:
Рис. 11.21 Эффекты действия ПТГ: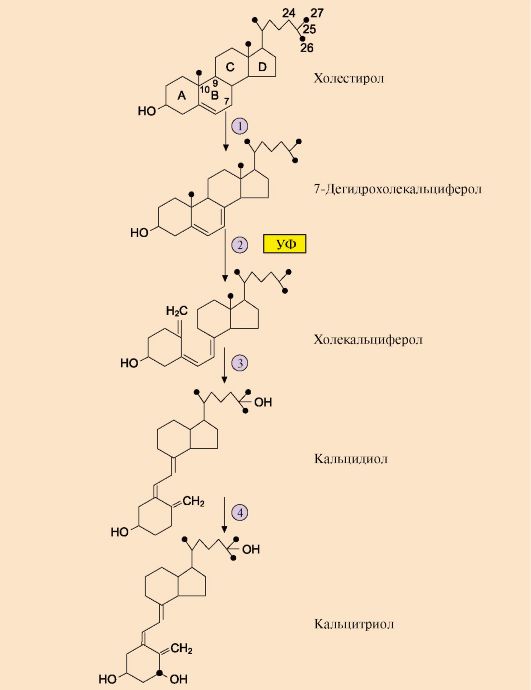 Рис. 11.22 Схема синтеза кальцитриола:
Рис. 11.22 Схема синтеза кальцитриола: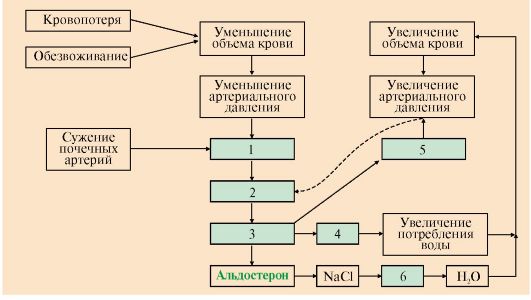 Рис. 11.23. Схема регуляции водно-солевого гомеостаза
Рис. 11.23. Схема регуляции водно-солевого гомеостаза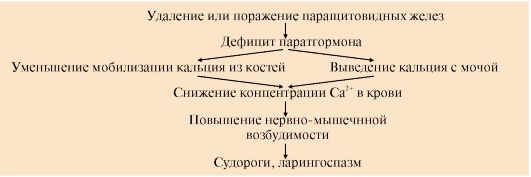
 ВКонтакте
ВКонтакте Facebook
Facebook Twitter
Twitter